HARVARD PAPERS IN BOTANY 5(1): 113-121. 2000.
IHSAN A. AL-SHEHBAZ(1), KATSUTOSHI ARAI(2) AND HIDEAKI OHBA(3)
(1)Missouri Botanical Garden, P.O. Box 299, St. Louis, Missouri 63166-0299, U.S.A.
(2)Karuizawa Botanical Garden, Hocchi 1166, Karuizawa, Kitasaku-gun, Nagano Pref., Japan.
(3)Department of Botany, University Museum, University of Tokyo, 7-3-1 Hongo, Tokyo, 1 13 Japan.
Abstract. Four species of Lignariella are recognized, of which two, Lignariella hinkuensis and L. ohbana, are described as new. The new combination, L. serpens, is proposed. The generic limits and relatives of Lignariella are discussed, and a key to related genera is provided.
Keywords: Brassicaceae. Cochlearia, Lignariella, taxonomy.
The genus Lignariella Baehni was established by Baehni (1955) on the basis of two species described by Pearson (1900) and Smith (in Smith and Cave, 1911) as Cochlearia hobsonii Pearson and C. serpens W. W. Smith, respectively, though both Pearson and Smith indicated that their species are anomalous in Cochlearia and should perhaps be accommodated in a different genus. Baehni provided a detailed account of Lignariella and the justification to separate it from Cochlearia. We fully support his position on the generic lines, but we disagree with his reduction of C. serpens to synonymy of L. hobsonii, as the two species differ drastically in flower size and leaf morphology (see below). The characters that readily separate Lignariella from the unrelated Cochlearia include purple to lavender petals, palmately lobed leaves, fully bracteate racemes, veinless valves, eseptate fruits, and flattened replum. Cochlearia has white petals, unlobed leaves, ebracteate racemes, strongly veined valves, septate fruits, and terete replum.
In his transfer of Draba obscura Dunn to Lignariella, Jafri (1957) confused the generic limits of Lignariella and recognized as conspecific two species of two entirely different genera. The type of D. obscura is definitely a member of the genus Aphragmus Andrzejowski ex de Candolle, and Schulz (1933) had correctly transferred it to that genus, though he (Schulz, 1926) described it originally as A. himalaicus O. E. Schulz but corrected that in his 1933 publication. All except three collections that Jafri (1957) cited under L. obscura belong to A. obscurus (Dunn) O. E. Schulz. The other three collections are assigned herein (see below) to the new species L. ohbana.
Hara (1975) was the first to notice this mixup of Jafri's (1957) species and generic concepts, and he indicated that "on examination of the type of Draba obscura Dunn at Kew, I noticed, contrary to expectation, that Aphragmus obscurus (Dunn) O. E. Schulz is quite a different plant from Lignariella" (p. 44). However, Hara treated L. serpens as a subspecies of L. hobsonii instead of as an independent species.
Based on the examination type, authentic, and numerous other collections of all species of Aphragmus and Lignariella, we conclude that both genera are well marked and should be recognized as independent. Aphragmus consists of rhizomatous or cespitose perennials with white or creamy white petals, long filiform funicles, entire, pinnately veined leaves, poorly developed nectaries often with deficient or obsolete median glands, unequal filaments not dilated at base, and sessile fruits. By contrast, species of Lignariella are biennials or short-lived perennials with slender taproots terminated by remains of basal rosettes, and they have purple petals, very short and thick funicles, 3-, 5-, or 7-lobed, palmately veined leaves, well-developed nectaries with confluent median glands, equal filaments dilated at base, and gynophorate fruits. Both genera have eseptate fruits with slightly flattened repla and minutely papillate seeds, and they are somewhat closely related.
Naqshi (1983) described Lignariella duthiei Naqshi from Kashmir and indicated that the plant is cespitose with woody radix, rosulate basal leaves, branched hairs, pinnate leaf venation (see illustration), and pedicels 2-3 mm long. None of these features are found in Lignariella, and all are typical characteristics of the Himalayan Braya. Hajra et al. (1993) were correct in reducing the species to synonymy of Aphragmus oxycarpus (J. D. Hooker & Thompson) Jafri sensu Jafri (1956).
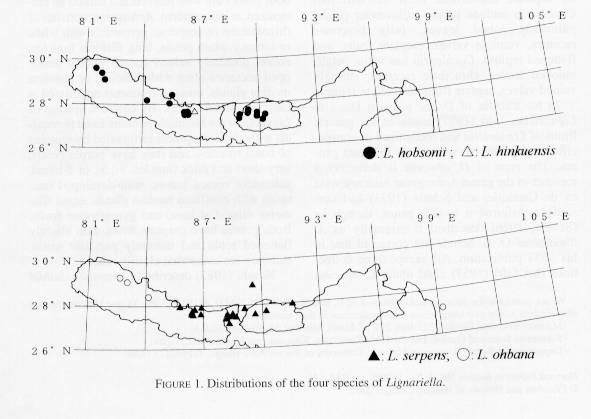
Perhaps Lignariella (Fig. 1) is most closely related to Himalayan-Central Asian (China, Kazakstan, Kyrgyzstan, Mongolia, Tajikistan) Taphrospermum C. A. Meyer (seven species) and the Chinese Platycraspedum O. E. Schulz (two species). All three have racemes bracteate throughout, leafy bracts, flattened replum, filaments dilated at base, and simple, eglandular, papillate trichomes. From these genera, Lignariella is readily separated by having purple to lavender flowers, palmately 3-, 5-, or 7-lobed leaves, spreading filaments equal in length, nonfleshy taproots, pedicels papillate along adaxial line, veinless valves, and eseptate fruits. By contrast, Taphrospermum and Platycraspedum have white flowers, unlobed leaves, fleshy taproots, glabrous or uniformly pubescent pedicels, veined valves, and septate fruits.
Schulz (1936) recognized both Lignariella hobsonii and L. serpens as distinct species of Cochlearia sect. Hilliella O. E. Schulz. The section was reduced by Al-Shehbaz et al. (1998) to synonymy of the Chinese endemic Yinshania Ma and Y. Z. Zhao, and neither species was treated in that genus. In their placement of Lignariella in the tribe Lepidieae, Baehni (1955) and Kuan (1987) followed Schulz (1936) in assigning Cochlearia and its segregates to that tribe. However, a closer examination of the flowers, fruit, replum, and leaves of Lignariella clearly support a closer association with Cardamine L. (tribe Arabideae), especially its former segregate Loxostemon J. D. Hooker & Thomson, which also has purple to lavender flowers, palmately lobed leaves, stamens somewhat equal in length, and flattened replum. It is interesting to note that Schulz (1936) placed Taphrospermum in the tribe Sisymbrieae and Platycraspedum and Cochlearia in the Lepidieae, and none of these genera were treated in the Arabideae.
There is a striking resemblance between Gignariella and some species of Eutrerrea R. Br., especially E. tenuis (Miquel) Koidzumi, in having inflorescences bracteate throughout, leafy bracts, palmately veined leaves, confluent nectar glands, dilated filament bases, and shortstipitatefruits. However, Lignariella differs in having nonfleshy taproots, palmately lobed leaves, purple to lavender petals, spreading stamens equal in length, flattened replum, and eseptate fruits. By contrast, the palmately veined species of Eutrema have fleshy roots, unlobed leaves, white petals, erect stamens unequal in length, terete replum, and septate fruits.
KEY TO THE GENERA ALLIED TO LIGNARIELLA
1a. Racemes ebracteate or only lowermost few dowers bracteate … 2
1b. Racemes bracteate throughout … 5
2a. Fruit a linear silique, flattened parallel to septum, dehiscing explosively; replum flattened; valves coiled during dehiscence ... Cardamine
2b. Fruit an oblong to elliptic of ovoid to obovoid silicle, rarely linear, terete, not dehiscing explosively; replum rounded: valves not coiled during dehiscence ... 3
3a. Septum absent; funicles long and filiform; lowermost flowers bracteate; basal leaves subsessile or short petiolate ... Aphragmus
3b. Septum complete or perforated: funicles short and stout; lowermost flowers ebracteate or, if bracteate, then leaves palmately veined; basal leaves often long petiolate ...
4a. Fruit valves strongly veined; seeds biseriate; median nectar glands absent; cotyledons accumbent; cauline leaves often fleshy ... Cochlearia
4b. Fruit valves not or obscurely veined; seeds uniseriate; median nectar glands present; cotyledons incumbent … Eutrema
5a. Petals purple to lavender; middle and upper leaves 3-, 5-, or 7-lobed; filaments spreading, equal in length; fruit stipitate or rarely subsessile; pedicels papillate adaxially ... Lignariella
5b. Petals white; middle and upper leaves not lobed; filaments erect to erect-ascending, unequal in length; fruits sessile or rarely short- stipitate; pedicels glabrous or usually evenly pubescent ... 6
6a. Leaves pinnately veined; septum absent; funicles long and tiliform; taproots not fleshy; seeds biserrate … Aphragmus
6b. Leaves palmately veined (pinnate in Taphrospermum); septum complete or perforate; funicles short and thick; taproots usually fleshy; seeds uniseriate … 7
7a. Median filaments strongly flattened, laterally 1-toothed; lateral sepals saccate; median nectaries absent; stigma 2-lobed ... Platycraspedum
7b. Median filaments terete, toothless; lateral sepals not saccate; median nectaries present; stigma entire or subentire … 8
8a. Leaves palmately veined; replum terete; seeds reticulate … Eutrema
8b. Leaves pinnately veined; replum flattened; seeds foveolate or papillate … Taphrospermum
TAXONOMIC TREATMENT OF LIGNARIELLA
Lignariella Baehni, Candollea 15: 57. 1955.
Type species: L. hobsonii (Pearson) Baehni.
Herbs biennial or short-lived perennial. Trichomes unicellular, eglandular, simple, clavate papillae. Stems ascending or decumbent to prostrate, papillate or glabrous, 1 to many originating from center of scalelike basal leaf rosette of previous year(s). Basal leaves not seen. Cauline leaves petiolate, palmately 3(-5)lobed or 3(-5)-partite to base, sometimes 3(-5)toothed near apex, rarely repand or entire, glabrous, usually palmately veined. Racemes corymbose, several-flowered, bracteate throughout, terminal, elongated considerably in fruit; rachis straight; bracts similar to cauline leaves. Fruiting pedicels slender, terete, divaricate and often recurved or reflexed, rarely straight, papillate adaxially, persistent. Sepals oblong, free, deciduous, ascending-spreading, glabrous, equal, bases of all subsaccate, rarely not saccate, margin membranous. Petals purple, blue, or pale lavender, spreading, often much longer than sepals; blade orbicular to narrowly obovate, rounded at apex; claw very short. Stamens 6, spreading, equal in length; filaments filiform, unappendaged, distinctly dilated at base; anthers narrowly oblong to ovate, apiculate or not at apex. Nectar glands confluent and subtending bases of all stamens; median nectaries present; lateral nectaries semiannular. Ovules 2-6(-12) per ovary. Fruit dehiscent siliques or silicles, linear, oblong, ovoid, or ellipsoid, terete or slightly latiseptate, not inflated, subsessile or long stipitate; valves thin, papery, veinless, glabrous or sparsely papillate, smooth or slightly torulose; gynophore short and to 1 mm long, sometimes 2-10 cm long; replum slightly flattened; septum absent; style short or to 4 mm long, sometimes recurved; stigma capitate, entire. Seeds uniseriate, rarely 1, wingless, oblong, plump; seed coat minutely colliculate, not mucilaginous when wetted; cotyledons incumbent.
A genus of four species distributed in Bhutan, China, India, Nepal, and Sikkim.
KEY TO THE SPECIES OF LIGNARIELLA
la. Fruit 1-seeded; gynophore 0.5-1.0 mm in flower, 20-100 mm in fruit; seed 3.7-4.7 mm long ... 2. L. hinkuensis
lb. Fruit 2- to 8(-12)-seeded; gynophore obsolete in flower and fruit, rarely to 2 mm in fruit; seed to 2.6 mm long ... 2
2a. Petals orbicular or orbicular-obovate, (5.5-)6.0-8.0(-9.0) mm long; anthers 0.6-1.2 mm long ... 1. L. hobsonii
2b. Petals narrowly to broadly obovate, 1.5-4.0(-4.5) mm long; anthers 0.25-0.40 mm long … 3
3a. Fruit oblong-ovoid to broadly oblong 2- to 4-seeded; fruiting pedicels often recurved; petals purple, (2.7-)3.0-4.0(-4.5) mm long, 1.7-2.5 mm wide; all stem leaves 3- or 5(or 7)-lobed ... 3. L. serpens
3b. Fruit narrowly oblong to linear, 6-12 seeded; fruiting pedicels usually straight; petals pale lavender, 1.5-2.0 mm long, 0.5-1.0 mm wide; lower stem leaves simple … 4. L. ohbana
1. Lignariella hobsonii (H. Pearson) Baehni, Candollea 15: 57. 1955.
Cochlearia hobsonii H. Pearson, Hook. Icon. Pl. 7: t. 2643. 1900. TYPE: TIBET. Yatung, near the Sikkim border, [27° 51' N, 88° 35' E, 1897], Hobson s.n. (Holotype: K).
Plants short-lived perennial, puberulent on stem and/or pedicels. Stems coarse, few to many from base, decumbent or rarely ascending, (6-)11-20(-30) cm long, glabrous or puberulent along 1 line or throughout. Leaves simple or rarely deeply 3(or 5)-lobed apically; petiole somewhat flattened, 4-10(-18) mm long; leaf blade broadly obovate to suborbicular and often wider than long, rarely oblong, (3-)6-10(-15) mm long, (1.5-)4.0-10.0(-15.0) mm wide, glabrous, obscurely to prominently 3or 5-toothed, teeth sometimes restricted to subtruncate apex, when lobed central lobe broadly to narrowly oblong, slightly broader than lateral lobes. Flowers often protogynous. Fruiting pedicels slender, strongly recurved to sigmoid, (0.7-)1.5-3.00.0) cm long, adaxially puberulent with papillae to 0.1 mm long. Sepals oblong, glabrous, 2.5-3.5(-4.0) mm long, 1.0-1.5 mm wide, margin membranous and to 0.5 mm wide, spreading, slightly saccate at base. Petals blue to deep purple, orbicular to orbicular-obovate, (5.5-)6.0-8.0(-9.0) mm long, 4-6(-7) mm wide, apex rounded, base abruptly narrowed into a claw 0.5-1.5 mm long. Filaments purple distally, whitish basally, 4.0-5.5 mm long; anthers narrowly oblong, 0.6-1.2 mm long, often strongly curved after dehiscence. Ovary 4- to 6(to 11)-ovuled. Fruit linear to narrowly oblong, rarely ovoid, not geocarpic, straight or curved, (5-)10-20 x 1.5-2.0 mm; gynophore ca. 0.5 mm long; valves torulose; style 3-4 mm long, stout. Seeds uniseriate, oblong, 2.2-2.8 x 1.2-1.4 mm.
Phenology: flowering May-July, fruiting June-September.
Altitude: 2800-4100 m.
Habitats: river or stream banks on small pebbles, girt, wet grass streamside, mossy area in running water, wet cliffs, scree, damp banks, wet stony slopes.
Specimens examined: BHUTAN. Barshong, Nala, H. Kanai, G. Murata, H. Ohashi, O. Tanaka, & T. Yamazaki 6045 (MO, TI); Barshong, Nala, H. Kanai, G. Murata, H. Ohashi, O. Tanaka, & T. Yamazaki 6018 (TI); Barshong, Shodu, H. Kanai, G. Murata, H. Ohashi, O. Tanaka, & T. Yamazaki 6791(TI); below Barshong, Thimbu Chu, Ludlow, Sherriff, & Hicks 16365 (BM, E, KATH, TI); Zado La Limpa, Cooper 2735 (BM, E); Joedowachi, Cooper 4821 (BM, E); Dungshinggang, Ludlow & Sherriff 3085 (BM, KATH); Dunshingang ridge, Dungchelakk, Bowes Lyon 3234 (BM); Thimbu Chu, Bowes Lyon 9019 (E); Zepla, Gould 1293 (BM); Thimpu District, Thimphu Chu, below Barshong, Wood 7295 (E). CHINA. Xizang (Tibet): Yaidong, Zhong Bubeng 5983 (PE); Yaidong Doggela Shan, Qinghai-Xizang Expedition 750248 (HNWP, KUN, PE); Nelamu Xian, Qinghai-Xizang Expedition 6139 (KUN, PE). NEPAL. Lantang, Polunin 466 (BM); Tori Dwari, Shrestha 5255 (BM, TI); Rambrong, Lamjung Himal, Stainton, Sykes & Williams 6017 (A, BM, E, UPS); Bhurchula Lekh, near Jumla, Polunin, Sykes & Williams 4610 (BM, E); Khimti Khola, 86° 17' E, 27° 38' N, Stainton 4785 (BM, E, TI); Maulekh, Stainton 8359 (E); Jumbesi, Stainton 7175 (BM, TI); Kalinchock, Dolakha District, Sharma, Subedi, & Kurmi 34 (KATH); Sagarmatha Zone, Solukhumbu Distr., GnaulaPike, Miyamoto et al. 9592074 (TI); Pike-Pike Peak, Miyamoto et al. 9592087 (TI).
Of the three sheets at Kew that have Hobson's collection label from Yatung (Tibet), the one bearing "Cochlearia hobsonii Pearson Hook. Ic. t. 2643" and the pencilled drawing on which the plate in Pearson (1900) was based is the holotype. This sheet, which was annotated by Baehni in 1954 as "type du Cochlearia hobsoni Pearson," has 12 fragments of one species with fruiting styles 2.5- 4.0 mm long, anthers 0.60-0.75 mm long, unlobed bracts, and orbicular petals ca. 5 x 5 mm.
The second sheet has two packets and a mounted small plant. It has the printed label "Field No. 5795 Herb. G. Watt" and the handwritten collection data "Jongri, 14,000 ft., Sikkim, Aug. 1881, ex Herb G. Watt, March 1881:" This sheet was cited by Smith and Cave (1911), and it is the syntype of Cochlearia serpens W. W. Smith. The plants on this sheet have fruiting styles 0.5-1.0 mm long, anthers 0.25-0.40 mm long, deeply 3-lobed bracts, and obovate petals 3.0-3.5 x ca. 2.0 mm. It is designated by us as the lectotype of C. serpens (see discussion under Lignariella serpens).
The third sheet, which was also annotated by Baehni as the type of Lignariella hobsonii, includes two species, of which one is represented by six fragments of L. hobsonii and the other by two of L. serpens. This sheet is not considered in the typification of either species of Lignariella. Baehni (1955), Jafri (1957), and others who studied the three Hobson's collections at Kew overlooked the fact that two species were involved. Because of this and because of Jafri's (1957) confusion between the identity of C. serpens and Draba obscura (see above), the limits within Lignariella were confused for more than 40 years.
Although Pearson's (1900) illustration shows that the fruits of Lignariella hobsonii are oblong-ovoid, in the majority of specimens examined they are linear to linear-oblong, and rarely are they similar to those of the type collection. The number of ovules maturing into seeds varies greatly, and the fruit shape appears to be correlated with the number of mature seeds. The species predominantly has 3- or 5toothed leaves apically, and rarely are the leaves typically 3- or 5-lobed. The later case is typical of plants of L. serpens.
Both Pearson (1900) and Baehni (1955) reported glandular hairs on the pedicels of Gignariella hobsonii. A close examination of these clearly reveals that they are clavate papillae, much like those common in the rest of the genus and in many species of Lepidium. True glandular hairs make the plant sticky, and they are found in many Central Asian genera, such Chorispora R. Brown ex DC., Dontostemon Andrzejowski ex DC., Hesperis L., and Sterigmostemum Poiret, as well as in various New World species of Descurainia Webb & Berthelot.
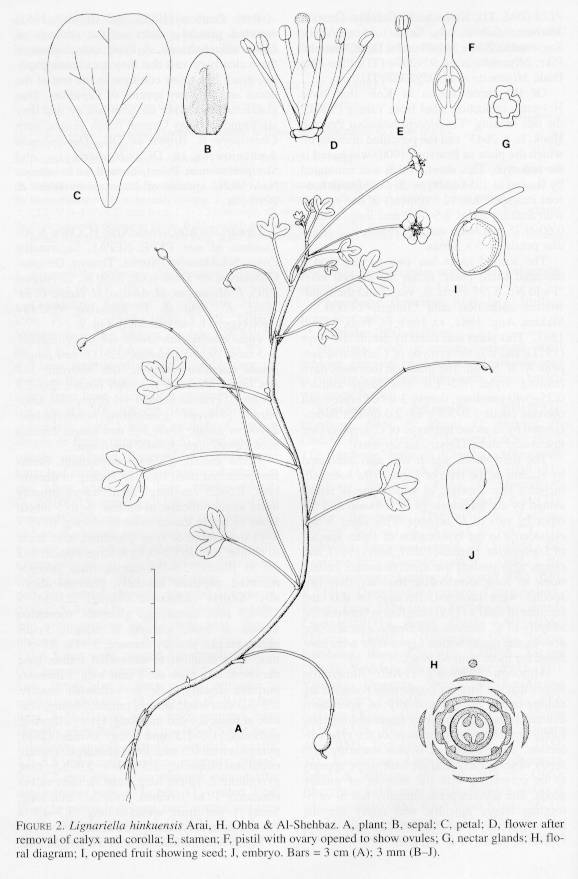
2. Lignariella hinkuensis Arai, H. Ohba & Al-Shehbaz, sp. nov. TYPE: NEPAL. Sagarmatha Zone: Solukhumbu District, Thasing DingmaChhatarwa, on a wet rock, 3550 m, 28 August 1995, F. Miyamoto, M. Amano, H. Ikeda, C. M. Joshi, K. Arai & T. Komatsu 9592484 (Holotype: TI; Isotype: MO). Fig. 2.
Folia caulina late ovata vet suborbiculata, 3-5 loba. Sepals oblonga, 2.3-3.0 mm Tonga; petala caeruleo-purpurea, late obovata, 5-7 mm Tonga. Pedicelli fructiferi tenues, 2.0-7.5 cm longi. Fructus ovoidei vet ellipsoidei, unispermi, geotropi; gynophoro tenuississimo, 2-10 cm longo; stylo 2-3 mm longo. Semina ovoideo-oblonga, 3.7–1.7 mm longa.
Herbs annual, glabrous throughout. Stems few-branched from base, ascending to decumbent, 8.5-25 cm long. Stem leaves broadly ovate to suborbicular in outline, 3- to 5-lobed; lobes of larger leaves ovate to oblong, 6-15 x 4-11 mm, entire or 1- or 2-toothed, apex acute to obtuse; petiole 1.5-3.7 cm long. Pedicel 1-2 cm in flower, 2.0-7.5 em in fruit, strongly recurved, papillate adaxially, glabrous abaxially. Sepals caducous, oblong, 2.3-3.0 x 1.2-1.7 mm, ascending, glabrous, somewhat saccate at base, scarious at margin. Petals bluish purple, broadly obovate, 5 -7 x 2.7-5.8 mm, apex rounded to somewhat retuse, base narrowed to a claw ca. 1 mm long. Filaments purplish distally, pale to yellowish basally, 2.5-3.0 mm long; anthers purple, oblong, cordate at base, 0.4-0.6 mm long. Ovary ellipsoid, glabrous, 1.0-1.5 mm long; ovules (2-)4; gynophore ca. 0.5 mm. Fruit geocarpic, curved, ovoid to ellipsoid, 2.5-7.0 x 2.0-5.5 mm; gynophore 2-10 cm long, very slender; valves obscurely 1- to 3-veined; style 2-3 mm long. Seeds 1 per fruit, ovoid-oblong, 3.7.7 x 1.0-1.2 mm; cotyledons incumbent.
Lignariella hinkuensis, which is named after the Hinku River (Nepal) along which the type was collected, is known thus far only from the type gathering. It is readily distinguished from L. hobsonii and L. serpens by the much larger (3.7-4.7 mm instead of 1.0-2.8 mm) seed size, 1-seeded instead of 2- to 12-seeded fruits, longer (2-10 cm instead of 0.2 cm or less) gynophores in fruit, and development of welldefined geocarpy. In L. hobsonii and L. serpens the fruiting pedicels are sometimes recurved and show a tendency toward geocarpy. It is safe to conclude that in L. hinkuensis the reduction of seed number, increase in seed size, and development of long gynophores and geocarpy are advanced characters in Lignariella.
The development in Lignariella hinkuensis of gynophores to 10 cm long is rather remarkable, because in no other genus in the entire Brassicaceae do the gynophores reach that length. The previous records of longest gynophores of 2.5-4.0 cm are known from the European Lunaria rediviva L. (Ball, 1993), of 2.0-3.2 cm in Romanschulziu mexicana Iltis & Al-Shehbaz (Al-Shehbaz and Iltis, 1993), and of 1.0-2.5 cm in the western U.S. Stanleya pinnata (Pursh) Britton and S. viridiflora Nuttall ex Torrey & A. Gray (Rollins, 1993).
The other remarkable feature of Lignariella hinkuensis is the development of geocarpy. Although the fruiting pedicels in L. serpens and L. hobsonii are sometimes strongly recurved, it is the combination of long pedicels and gynophores that help in planting the one-seeded fruits at some distance from the mother plant. This is the first report of geocarpy in any Asian species of the Brassicaceae. Geocarpy is well documented in the South American Cardamine chenopodiifolia Persoon, the Australian Geococcus pusillus Drummond ex Harvey, and the Mediterranean (Corsica, Sardinia) Morisia monanthos (Viviani) Ascherson (Al-Shehbaz, 1984; Schulz, 1919, 1936; Shaw, 1965). All these unrelated species, including L. hinkuensis, have indehiscent or tardily dehiscent, one- or few-seeded fruits. Evidently, geocarpy evolved independently at least four times in the family. Fruits of Northwest African (Algeria and Morocco) Raffenaldia Godron (two species) show strong tendencies to geocarpy, and critical field work is needed to establish that.
3. Lignariella serpens (W. W. Smith) AlShehbaz, Arai, & H. Ohba, comb. nov.
Cochlearia ?serpens W. W. Smith in W. W. Smith & G. H. Cave, Record. Bot. Surv. India 4: 175. 1911. TYPE: SIKKIM HIMALAYA. Jongri, 14,000 ft, Watt 5795 (Lectotype, here designated: K).
Plants slender, short-lived perennial, glabrous throughout except for pedicels and sometimes fruit. Stems slender, few to many from base, decumbent, (I.5-)5.0-15.0(-30.0) cm long. Leaves deeply 3(or 5)-lobed throughout, simple leaves absent; petiole glabrous, (2-)5-1500) mm long; blade of central lobe oblong to ovate, rarely linear, entire or rarely 1toothed on each side, (1.5-)3.0-8.0(-15.0) mm long, (0.5-)1.0-3.0(-7.0) mm wide, apex obtuse to subacute; lateral lobes smaller, entire, 1- or 2-lobed or toothed , rarely leaves subternate. Fruiting pedicel (10-)15-3000) mm long, strongly recurved to sigmoid, often filiform, adaxially puberulent with papillae to 0.1 mm long. Sepals oblong, 1.5-5.0 x 0.7-1.0 mm, not saccate at base, scarious margin ca. 0.2 mm wide. Petals purple, broadly obovate, (2.0 )3.0-4.0(-4.5) mm long, (1.0-j1.5-2.5(-3.0) mm wide, apex rounded, cuneate into a claw to 0.1 mm long. Filaments purplish, 1.5-2.5 mm long; anthers purple, ovate, 0.3-0.4 mm long. Ovary 3- to 6-ovuled. Fruit ovoid to oblong, rarely oblong-linear, not geocarpic, often curved at middle, 2.5-8.0(-12.0) x 1.0-2.0(-2.5) mm; gynophore 0.5-3.0 mm long; valves slightly torulose or not, glabrous or puberulent; style 1.0-1.5(-2.0) mm long, stout. Seeds uniseriate, oblong, (1.0-)1.5-2.5 x (0.5-)0.8-1.2(-1.5) mm.
Phenology: flowering June-August, fruiting July-October.
Habitat: alpine peaty soil, scree, gravelly stream edges, turf among rocks and small shrubs.
Altitudes: 2600–4300 m.
Specimens examined: BHUTAN. Ju La, Mangde Chu, Ludlow & Sherriff 16897 (BM). CHINA. Gyanke, Benham s.n. (BM); Tare-La, above Ha, Could /203 (K); Shingbe (Me La); Ludlow & Sherriff 20318 (BM), Ludlow, Sherriff & Hicks 20714 (BM); Thimphu District, near Darkey Pang Tso, N of Paro, Wood 7397 (E). NEPAL. Rolwaling, 86° 28' E, 27° 55' N, Stainton 4675 (E, BM, TI); Chairam, Williams 890 (TI); Sagarmatha Zone, Solukhumbu District, Pike-Pike Peak, Miamoto et al. 9592090 (TI); Solukhumbu District, Chalem Kharka-Jar Kharka, Miyamoto et al. 9592152 (TI); Bhararate Himal: Barn Valley, Yangle, Wraber 331 (BM); Khimti Khola, Bowes Lyon 2207, 2208 (BM); Likhu Khola, E of Chaukharma, 86° 15' E, 27° 30' N, McCo.sh 248 (BM, E); Bheging, Lall Dhwoj 263 (BM, E); Bhairavkunda, Maine 99 (BM). SIKKIM. Darjeeling, along Choktsering Chu, Hara et al. 6174 (TI); Gamotang, Hara et al. 6172, 6173 (TI); Changu, Smith 3577 (E), Smith 3061 (E, K), Smith 3109 (E), Cooper 9 (BM, E), Cooper 33 (BM, E), Cooper 846 (A, BM, E), 20 Sept. 1916, Cave s.n. (E), Ribu & Rohmoo 4141 (E); Jongre, 1 Sept. 1919, Cave s.n. (E), Ribu & Rohmoo 6566 (E); Yampung, Ribu & Rohmoo 1107 (E); Chumighata, Rohmoo Lepcha 538 (E); Dzongri, Staling, Upward, Brickell & Mathew 80 (E, K); Tha moo tsa, near Jongri, June 1887, King s.n. (K); Koteh ya rong, June 1888, King s.n. (E); Piamphomg, W Jongre, July 1887, King s.n. (K); Zemu Valley, Smith & Cave 1544 (K); Eermtso La, Smith & Cave 1269 (K); Dobiuda Pass, Cooper 340 (BM); Yampung, Ribu & Rohmoo 875 (E); West District, Bikbari, E slope of Choktsering Chhu valley, 88° 08' E, 27° 30' N, Long, McBeath, Noltie, & Watson 319 (E, TI); West District: Jamlinghang, Chhurong Chhu valley below Dzongri, 88° 09' 14" E, 27° 28' 20" N, Long, McBeath, Noltie, & Watson 196 (E, TI); Kapook, Rohmoo Lepcha 24 (E, K); Gamotang, Hara, Kanai, Murata, Togashi & Tuyama (BM, TI).
Three collections-Ludlow & Sherriff 16897 (BM), Ribu & Rohmoo 1107 (E), and Smith & Cave 1544 (K)-were cited by Baehni (1955) as L. hobsonii.
Although Smith (Smith and Cave, 1911) did not designate a holotype and cited four syntypes, Watt's specimen-Watt 5795 (K)which was accompanied by Smith's line drawings of a petal, stamen, and pistil, along with notes on these and other parts, is designated herein as the lectotype (see also discussion under Lignariella hobsonii). Two other syntypes, Smith and Cave 1544 and Smith & Cave 1269 (both at K), bear Smith's pencilled handwriting as "Cochlearia serpens W. W. Smith" and "=Watt 5795." The fourth syntype (Gammie No. 156) has not been found.
Lignariella serpens exhibits a tremendous variation in fruit morphology. As in L. hobsonii, the degree of fruit curvature depends on the development of seeds on one or both sides of the replum, and therefore, in strongly curved fruits, the ovules do not mature into seeds on one side of the fruit. This is well exhibited in Cooper 846 (BM).
Although confused with Lignariella hobsonii by Baehni (1955) and Jafri (1957), L. serpens has much smaller flowers (see key), and of the 94 specimens of the two species studied by us, the differences between the two species in petal size and anther length are consistent throughout. Furthermore, the leaves of L. serpens are never simple nor are they apically 3or 5-toothed, as in L. hobsonii. From L. ohbana, which has much shorter pedicels, L. serpens is readily separated by the total lack of simple, undivided lowermost leaves, which are so characteristic of L. ohbana.
4. Lignariella ohbana Al-Shehbaz & Arai, sp. nov. TYPE: NEPAL. Rambrong, Lamjung Himal, open slopes, 13,000 ft, 3 July 1954, Stainton, Sykes & Williams 6075 (Holotype: BM; Isotypes: A, E, UPS).
Folia caulina inferiors simplicia, .superiors 3-lobata. Sepala oblonga, 0.7-1.5 mm longa; petala la vandula, anguste obovata, 1.5-2.0 mm longa. Pedicelli fructiferi 5-8(-13) mm longi. Fructus lineari vel anguste oblongi, 6 ad 12spermi; gynophoro obsoleto; stylo 0.5-0.8 mm longo. Semina oblonga, 1-1.4 mm longa.
Plants slender, biennial or short-lived perennial, glabrous throughout except for pedicels. Stems slender, few or rarely 1 from base, decumbent, 2-12(-15) cm long. Lowermost stem leaves simple, becoming 3-lobed in the middle and distal portions of plant; petiole glabrous, (2-)4-8(-12) mm long; blade of lowermost leaves ovate, subacute, 3-5 mm long, 2-3 mm wide; middle lobe of bracts narrowly oblong to broadly ovate, entire, (2-)3-6(-8) mm long, 1.3-1.0(-7.0) mm wide, obtuse; lateral lobes smaller and narrower, entire or rarely obscurely 1-toothed. Fruiting pedicel 5-8(-13) mm long, adaxially puberulent with papillae to 0.1 mm long. Sepals oblong, 0.7-1.5 x 0.5-0.7 mm, not saccate at base, scarious at margin. Petals pale lavender, narrowly obovate, 1.5-2.0 mm long, 0.5-1.0 mm wide, apex rounded, cuneate and not clawed at base. Filaments white or rarely lavender, 0.8-1.2 mm long; anthers purple, ovate, 0.2-0.3 mm long. Ovary 6- to 12-ovuled. Fruit linear to narrowly oblong, not geocarpic, straight or slightly curved at middle, 5-10 x 0.7-1.0 mm; gynophore obsolete; valves torulose; style 0.5-0.8 mm long, stout. Seeds uniseriate, oblong, 1.0-1.4 x 0.5-1.0 mm.
Phenology: flowering July-August, fruiting July-October.
Habitat: among rocks, sandy flats by streamside.
Altitudes: 3000–4500 m.
Paratypes: CHINA. Yunnan: Chungtien Plateau, Feng 1662 (KUN). NEPAL: Rambrong, Lamjung Himal, Stainton, Sykes & Williams 6131 (A, BM, E, UPS); Langtang Valley, Polunin 632 (BM); near Jangla Bhanjyang, Polunin, Sykes & Williams 2369 (BM, E); Lari, 88° 11' E, 28° 14' N, Yon 208 (BM). Maharigaon, Polunin, Sykes & Williams 295 (BM).
Lignariella ohbana is named in honor of our colleague Professor Hideaki Ohba for his outstanding contributions to Himalayan botany and for his recognition of the taxonomic difficulties in Lignariella that resulted in the supervision of a master's thesis by one of us (Arai, 1997). A photograph of the holotype of L. ohbana and a simple line drawing of the species are presented in Jafri (1957) as L. obscura. The label of the holotype was hand written, whereas that of the isotypes was typed, which resulted in the erroneous typing of the collection number as 6057 instead of 6075. All other data on the labels of the holotype and isotype sheets are identical.
Two of the paratypes above (Polunin et al. 295, 2369, both at BM) were cited by Baehni (1955) as Lignariella hobsonii. On the other hand only 3 (Polunin 632; Stainton, Sykes & Williams 6075, 6131) out of 13 collections cited by Jafri (1957) as Lignariella belong to L. ohbana, and the remaining 10 clearly belong to Aphragmus obscurus, a species that Jafri confused with the true members of Lignariella (see above). By contrast, none of the 12 collections cited by Jafri (1973) under Lignariella are members of this genus, as they belong to typical Aphragmus. Therefore, contrary to the reports of Jafri (1973), Hajra et al. (1993), and Srivastava (1998), the genus Lignariella does not occur in Pakistan or Kashmir. The disjunction between the single locality in Yunnan and the main distribution in Nepal is rather remarkable. It is unknown if this disjuction is the result of a recent introduction to China or lack of collections from the area in between.
ACKNOWLEDGMENTS
We are grateful to the directors and curators of A, B, BM, E, HNWP, K, KATH, KUN, MO, PE, TI, and UPS for the loan of specimens and/or providing access to the collections in their institutions.
LITERATURE CITED
AL-SHEHBAZ, I. A. 1984. The genera of Cruciferae (Brassicaceae) in the southeastern United States. J. Arnold Arbor. 65: 343-373.
AL-SHEHBAZ, 1. A., AND H. H. ILTIS. 1993. Romanschulzia mexicana (Brassicaceae), a remarkable new species from Guerrero, Mexico. Novon 3: 96-98.
AL-SHEHBAZ, 1. A., G. YANG, L. L. LL, AND T. Y. CHEO. 1998. Delimitation of the Chinese genera Yinshania, Hilliella, and Cochleariella (Brassicaceae). Harvard Pap. Bot. 3(1): 79-94.
ARAI, K. 1997. A taxonomic study of Lignariella (Cruciferae). M.S. thesis, Faculty of Agriculture, Tokyo University of Agriculture, Japan.
BAEHNI, C. 1955. Lignariella, genre nouveau de Cruciferes. Candollea 15: 47-62.
BALL, P. W. 1993. Luruzria. In T. G. TUTIN ET AL., eds., Flora Europaea, 2d ed., 1: 358. Cambridge University Press, Cambridge.
HAJRA, P. K., H. J. CHOWDHERY, AND G. C. DAS. 1993. Sisymbrieae. In B. D. SHARMA AND N. P. BALAKRISHNAN, eds., Flora of India 2: 224-247. Botanical Survey of India, Calcutta.
HARA, H. 1975. Cruciferae. Pages 43-45 in H. OHASHt, ed., The Flora of Eastern Himalaya. Tokyo University Press.
JAFRI, S. M. H. 1956. Some Cruciferae of W. Pakistan, Afghanistan and N.W. Himalaya. Notes Roy. Bot. Gard. Edinburgh 22: 95-1 19.
--. 1957. Another species of Lignariella Baehni. Candollea 16: 131-136.
--. 1973. Brassicaceae. In E. NASIR AND S. I. ALI, eds., Flora of Pakistan 55: I-308.
KLAN, K. C. 1987. Lepidieae. In T. Y. CHEO, ed., Cruciferae. Fl. Reipubl. Popularis Sin. 33: 44-109.
NAQSHI, A. R. 1983. On the Himalayan genus Lignariella (Brassicaceae). J. Econ. Tax. Bot. 3: 974-977.
PEARSON, H. H. W. 1900. Cochlearia hobsonii. Hook. Icon. Pl. 7: t. 2643.
ROLLINS, R. C. 1993. The Cruciferae of Continental North America. Stanford University Press, Stanford.
SCHULZ, O. E. 1919. Cruciferae-Brassiceae. Part 1. In A. ENGLER, ed., Pflanzenreich IV. 105(Heft 70): 1-290.
--. 1926. Beitrage zur Kenntnis der Cruciferen des nordwestlichen Himalaya-Gebriges. Notizbl. Bot. Gart. Berlin-Dahlem 9: 1057-1095.
--. 1933. Seltene Cruciferen aus Kashmir. Repert. Spec. Nov. Regni Veg. 31: 330-334.
--. 1936. Cruciferae. In A. ENGLER, ed., Naturl. Pflanzenfam. 1713: 227-568. Wilhelm Engelmann, Leipzig.
SHAW, E. A. 1965. Taxonomic revision of some Australian endemic genera of Cruciferae. Trans. Proc. Roy. Soc. S. Austral. 89: 145-253.
SMITH, W. W., AND G. H. Cave. 191 1. The vegetation of Zemu and Llonak valleys of Sikkim. Record. Bot. Surv. India 4: 141-260.
SRIvAsTAvA, R. C. 1998. Flora of Sikkim. Oriental Enterprises, Debra Dun.
{kind=link}
{kind=link}