4. Resultados y discusion
En este capítulo se presenta un análisis de los datos obtenidos en este estudio: la descripción de los tipos de vegetación en base de los géneros de leñosas registrados, la diversidad y se propone una clasificación fitogeográfica. Los resultados se comparan con aquellos obtenidos en otras zonas montañosas con vegetación similar.
Tipos de vegetación de regiones montañosas neotropicales.
Humboldt (1815) en su clasificación fitogeográfica de América tropical (entre 10 grados al norte y 10 grados al sur de la línea equinoccial) denominó Regio Frigida (Tierra fría) o de Quercuum, Winteræ y Escalloniæ, a la región ubicada desde los 2145 hasta los 4797 metros de altitud a la que subdividió en tres unidades: a (2145-3120 m) o subfrígida, b (3120-3705 m) o páramo y g o rocosa (3705-4797 m). Desde ese entonces, varias son las clasificaciones que se han propuesto para la vegetación neotropical. A continuación se citan algunos ejemplos relacionados a las zonas montañosas de varios países.
Cuatrecasas (1957), en su esquema de los tipos de vegetación de la provincia norte andina (los Andes al norte de la latitud 5°S), consideró que el bosque andino se extiende entre 2400-2500 y 3500-3800 metros y los páramos sobre los 3200 metros hasta el límite de las nieves. En relación a la vegetación montana de Colombia, el mismo autor (Cuatrecasas, 1958) propuso divisiones similares a las anteriormente expuestas: la selva andina o bosque andino sobre los 2400 metros hasta los 3800 metros y el páramo sobre los 3800 metros, aunque localmente desde los 3200 metros, hasta el nivel de las nieves; además incluyó los prados, que corresponden a las áreas montañosas deforestadas entre 2100 y 3000 metros. Apreciaciones más detalladas de la zonificación altitudinal, se han obtenido en los diferentes estudios realizados en base de levantamientos de vegetación, a lo largo de transectos altitudinales en Colombia: en la Cordillera Central (van der Hammen et al., 1983), en la Sierra Nevada de Santa Marta (van der Hammen y Ruiz, 1984), en la Sabana de Bogotá (Hooghiemstra, 1984).
Vareschi (1970) presentó una clasificación para la vegetación de los Andes de Venezuela entre 2300 y 5000 m de altitud. El autor dividió en 11 franjas denominadas con letras de A a K desde la cumbre hacia la base: A y B (5000-4800 m) son regiones nivales, C (4800-4700 m) es la región subnival, D (4700-4600) marca el límite superior de los frailejones entre E-I (4600-2800) se extiende la propia región de los páramos y entre I-K es la distribución principal de Podocarpus. A 4500 m es el límite superior del "chirrivital que es la vegetación de árboles que forman grupos sobre el límite del bosque cerrado; a 4200 m es el límite de Polylepis; a 3800 m es la límite inferior de las plantas en cojín; a 3200 m es el límite superior de las selvas; a 2800 m es el límite inferior de Espeletia, a 2300 m es el límite inferior de las selvas de Podocarpus. En la cordillera de la Costa en Venezuela son de interés los estudios de Huber (1986) en "Rancho Grande", y de Steyermark y Huber (1978) en el Parque Nacional "El Avila".
Para el Perú los clásicos estudios de la vegetación son los de Weberbauer (1911, 1936, 1945) quien dividió la vegetación en tres grandes conjuntos e incluyó en la región central montañosa a: las vertientes occidentales, los territorios alto-andinos y los valles interandinos. Toda esta zona fue dividida en doce unidades que a su vez fueron analizadas en subunidades detalladas, a las que hay que añadir también la Ceja andina, que forma parte del tercer gran conjunto de las vertientes orientales de los Andes. Estas subdivisiones indican la complejidad de la zona andina de ese país.
Por último, un estudio reciente en Costa Rica ubica al bosque montano alto entre 2400 y 3200 metros, al bosque subalpino entre 3200 y 3400 metros y al páramo desde los 3300 hasta los 3800 metros de altitud (Kappelle et al., 1991).
Tipos de vegetación de los Andes del Ecuador
Para la vegetación del Ecuador se han sugerido varias clasificaciones, unas más detalladas que otras. Con relación a la región andina, la vegetación ha sido clasificada de la siguiente manera:
- Sodiro (1874) consideró tres zonas de vegetación: zona templada o subtropical, entre 400 y 2800 metros; zona subandina entre 2800 y 3400 metros y zona andina (sobre 3400 metros).
- Diels (1937) dividió al Ecuador Central (Sierra o alto andino) en: nivel selvoso de la Cordillera Occidental; valles longitudinales de la Cordillera Occidental; nivel selvoso de la Cordillera Oriental; ceja andina; región interandina cultivada e improductiva; páramos andinos y nivel andino alto.
- Acosta Solís (1968b) propuso 10 formaciones vegetales sobre los 1800 metros de altitud. En la Cordillera Occidental el autor reconoce: bosque subandino (1800-2800 m), Ceja andina (2800-3200 m), páramos y pajonales (3200-4750 m) y faja gélida (4750-6310 m). Entre ambas cordilleras se encuentran: región interandina (2000-3000 m) y valles secos (1500-2000 m). En la Cordillera Oriental son las mismas divisiones, pero las altitudes varían: Ceja andina (2800-3300); páramos y pajonales (3300-4750) y faja gélida entre 4750-5840 metros.
- Harling (1979) consideró nueve tipos de vegetación sobre los 2500 metros de altitud: bosque nublado (ca. 2500-3400 m); vegetación de quebradas y pastizales del norte; vegetación arbustiva del sur; vegetación seca arbustiva del extremo sur; desiertos y semi-desiertos interandinos; pajonales (3400-4000 m); páramos arbustivos y de almohadillas (4000-4500 m) y páramos desérticos (sobre 4000-4500 m).
- Espinosa (1948, 1949) presentó un valioso estudio sobre la vegetación de la provincia de Loja en el sur del Ecuador.
Finalmente, cabe mencionar el mapa ecológico del Ecuador de acuerdo al sistema de las zonas de vida de Holdrige, en el que la Sierra incluye las formaciones montano bajas, montanas y subalpinas que suman un total de 12 (Cañadas Cruz, 1983).
De acuerdo a observaciones propias realizadas en el Ecuador, la cota de los 2300-2500 metros de altitud, es el límite "natural" de distribución máxima de varias familias, géneros y especies que hacia arriba desaparecen. Este límite se ha tomado para delimitar, en sentido amplio, dos tipos de vegetación: subandina y andina. Los límites de estos tipos de vegetación son evidentemente muy variables y dependen de factores orográficos locales como son la exposición de la cordillera, la influencia de los vientos, la precipitación, la humedad, el tipo de suelo, además del grado de intervención humana.
En el presente estudio se ha dividido la vegetación andina ubicada sobre los 2400 metros en bosque andino (montano), bosque altoandino (montano alto), bosque de Polylepis y páramos, que se discuten a continuación. Se presenta también una descripción de la vegetación de la zona de transición. Una clasificación detallada, tomando en cuenta todas las formas de vida, se describe en Jørgensen y Ulloa Ulloa (1994).
Vegetación de transición y de zonas secas - La zona de transición corresponde al límite superior del bosque subandino y límite inferior del bosque andino. En esta franja ubicada alrededor de los 2300-2500 metros de altitud (localmente hasta 2600 m), se encuentran varios géneros que se extienden desde el
 |
 |
 |
 |
El bosque subandino progresivamente da paso al bosque andino, y la composición florística y la fisionomía del bosque va cambiando con la altitud. Así, familias, géneros y especies característicos de las zonas bajas son reemplazados por otros taxa que son propios de zonas más frías.
Bosque andino - También denominado bosque montano, bosque nublado o selva andina corresponde a la faja de vegetación arbolada que reemplaza al bosque subandino y que se ubica sobre los 2400(-2600) y 3000 metros de altitud.
Los bosques andinos se encuentran en las vertientes de las cordilleras y están mejor desarrollados en la vertiente externa de la Cordillera Oriental. Se caracterizan por tener árboles medianos, entre 9-15(-30) metros de alto; los troncos están cubiertos por una densa vegetación epífita de musgos, bromelias, orquídeas, helechos, licopodios, líquenes, hepáticas y briofitas. Géneros de leñosas característicos de estos bosques son, entre otros: Aegiphila, Alnus, Brunellia, Ceroxylon, Cinchona, Freziera, Hedyosmum, Ilex, Meliosma, Miconia, Myrcianthes, Ocotea, Palicourea, Saurauia, Solanum, Tournefortia y Weinmannia. Son también característicos los helechos arborescentes especialmente de los géneros Cyathea y Dicksonia, y también "surales" del género Chusquea. En los valles altos interandinos, que en la actualidad son zonas agrícolas, se encuentran remanentes de bosques especialmente en las quebradas y montañas aisladas. Son características especies como: Ambrosia arborescens, Baccharis latifolia, Barnadesia arborea, Bocconia integrifolia, Coriaria ruscifolia, Dodonaea viscosa, Delostoma integrifolium, Euphorbia laurifolia, Juglans neotropica, Mimosa quitensis, Myrcianthes rhopaloides, Myrica pubescens y Tecoma stans, que probablemente habrían ocupado gran parte del callejón interandino hoy ocupado por cultivos, pastizales y plantaciones de Eucalyptus globulus.
Bosque altoandino. - También llamado bosque alto montano, corresponde a los bosques que se encuentran entre 3000 y 3400 m de altitud. Estos bosques limitan en la parte superior con los páramos y/o los bosques de Polylepis. Estos bosques se caracterizan por árboles pequeños de 6-9 m de alto, con troncos retorcidos, muchas veces recostados; los árboles con troncos múltiples son frecuentes y las epífitas cubren la mayoría de los troncos. Géneros leñosos característicos son Barnadesia, Sessea, Clethra, Columellia, Drimys, Escallonia, Gaiadendron, Gordonia, Gynoxys, Hesperomeles, Myrcianthes, Podocarpus, Prumnopitys, Styrax, Symplocos Vallea, y Weinmannia. La franja superior del bosque alto andino que limita con el páramo a menudo se denomina Ceja andina.
Bosque de Polylepis. - Estos bosques se encuentran entre 3400-4000 (-4350) m. Son bosques fragmentados y aparecen entremezclados con grandes manchones de pajonales e incluso se presentan a manera de "islotes" boscosos en medio de los pajonales. Están conformados por árboles pequeños de 3-7 metros de alto. Géneros característicos son Brachyotum, Buddleja, Ceratostema, Clethra, Columellia, Diplostephium, Disterigma, Escallonia, Gaiadendron, Geissanthus, Gynoxys, Hesperomeles, Ilex, Miconia, Myrteola, Oreopanax, Polylepis, Tristerix y Weinmannia. En esta zona son frecuentes los bambusales del género Neurolepis.
La distribución fragmentada de estos bosques correspondería a una antigua distribución más extensa, como ya lo evidenció Cuatrecasas (1958). Estudios realizados en el Ecuador proponen que: los fragmentos son residuos de una distribución más amplia y alta del bosque, que corresponderían al verdadero límite superior -4100 metros en la Cordillera Oriental y 4350 metros en la Cordillera Occidental-, y que, dichos bosques han sido parcialmente destruidos por el hombre (Lægaard, 1992). La controversia sobre el verdadero límite superior del bosque sigue en discusión. Varias hipótesis han tratado de explicar la presencia de estos fragmentos de acuerdo a características del suelo, humedad o topografía, que según Lægaard no son evidentes en el Ecuador. En la Cordillera Oriental del Perú, los bosques montanos fragmentados están ubicados entre 3500-3700 metros de altitud (Young y León, 1990); mientras que en la Cordillera Oriental colombiana reliquias de bosques macizos se han observado hasta 4000 (-4200) metros de altitud (Cuatrecasas, 1958).
Páramos.
- Corresponden al cinturón más alto de vegetación andina y se extienden hasta el
límite de las nieves. Esta inmensas áreas abiertas, y típicamente sin árboles,
se ubican en los lomos de las cordilleras, en promedio sobre los 3300 metros
(Acosta Solís, 1984); en la provincia de Loja se consideran páramos sobre los
2800 metros de altitud (Acosta Solís, 1984; Espinosa, 1948). Tradicionalmente
los páramos se dividen en: 1) subpáramo o páramo arbustivo, que es una zona
arbustiva de transición entre el bosque andino y el páramo propiamente dicho;
2) el páramo propiamente dicho o pajonal y 3) el superpáramo. La vegetación
paramuna se caracteriza por macollas de gramíneas, plantas arrosetadas
acaulescentes (Valeriana)
y caulescentes (Espeletia),
en almohadillas (Azorella)
y arbustos micrófilos enanos (por ejemplo
Chuquiraga y
Loricaria).
Arbustos de Baccharis,
Brachyotum,
Bucquetia,
Escallonia,
Hesperomeles y
Miconia que se encuentran en el
subpáramo se hallan también en los bosques adyacentes.
 |
 |
 |
 |
 |
 |
Los pajonales son quemados regularmente por el hombre. En estas áreas, numerosas plantas han desarrollado estrategias adaptativas para regenerarse luego de las quemas. Entre éstas se pueden mencionar los siguientes géneros de leñosas: Baccharis, Brachyotum, Chuquiraga, Chusquea, Espeletia, Fuchsia, Gynoxys, Hesperomeles, Miconia, Monnina, Pentacalia, Pernettya, Ribes y Vaccinium (Lægaard, 1992), y probablemente Columellia (Brizicky y Stern, 1969). Mientras tanto, varias leñosas que no pueden resistir el fuego sobreviven en los fragmentos de bosques antes mencionados. Debido a estas circunstancias se considera que los pajonales bajo 4100-4300 m, son una vegetación parcialmente secundaria creada y mantenida por el hombre usando el fuego como herramienta principal (Lægaard, 1992).
Diversidad florística
El bosque nublado andino tropical, pese a ocupar un área relativamente pequeña, tiene una variación ecológica extremadamente rica, en relación con las tierras bajas, y varias zonas de vida comprimidas (Gentry, 1989). La vegetación de las estribaciones de la cordillera parece ser la más rica y la mayor diversidad de especies se encuentra en los bosques premontanos al pie de las vertientes de los Andes (Gentry, 1982). Balslev (1988) calcula que en el Ecuador, entre 900 y 3000 metros se encuentra el 49% de las especies de plantas del país. La lista anotada de las plantas con semillas de los Andes ecuatorianos, sobre los 2400 metros de altitud en un área de cerca de 45.000 km 2, contiene 4430 especies (Jørgensen y Ulloa Ulloa, 1994), que corresponden al 22-27% del total de especies de plantas vasculares del país- estimado entre 16.500 (Balslev y Renner, 1989) y 20.000 (Gentry, 1977) especies. Esta cifra demuestra la riqueza de la flora andina si la comparamos con las 3100 especies de plantas con flores listadas (Renner et al., 1990) o inclusive con el estimado de por lo menos 4000 especies de plantas vasculares (Balslev y Renner, 1989) para la región amazónica del Ecuador, en un área dos veces mayor que la de la zona andina. Sin duda la alta diversidad florística de los Andes está en relación con la gran variación ecológica.
En el presente estudio se reportan 289 géneros pertenecientes a 93 familias y se listan 1568 especies de árboles y arbustos nativos registrados sobre los 2400 metros de altitud. De éstos, siete géneros y 14 especies son pteridofitos (helechos arborescentes de Blechnaceae, Cyatheaceae y Dicksoniaceae), tres géneros y seis especies son gimnospermas (Ephedraceae y Podocarpaceae), ocho géneros y 18 especies son monocotiledóneas (Arecaceae y Poaceae) y 271 géneros y 1530 especies son dicotiledóneas.
Del total de especies se encontró que: 334 son árboles, 1071 son
arbustos, 105 son arbustos-árboles y 58 son sufrútices (incluidos algunos trepadores,
lianoides y epífitos facultativos especialmente de Ericaceae).
| Familia | Géneros | % | Familia | Géneros | % |
|---|---|---|---|---|---|
| Asteraceae | 44 | 15,2 | Rosaceae | 7 | 2,4 |
| Ericaceae | 17 | 5,8 | Cyatheaceae | 5 | 1,7 |
| Solanaceae | 16 | 5,5 | Arecaceae | 5 | 1,7 |
| Rubiaceae | 14 | 4,8 | Flacourtiaceae | 5 | 1,7 |
| Melastomataceae | 13 | 4,5 | Lamiaceae | 5 | 1,7 |
| Euphorbiaceae | 8 | 2,7 | Lauraceae | 5 | 1,7 |
| Fabaceae | 7 | 2,4 | |||
| Myrtaceae | 7 | 2,4 | Otras familias | 131 | 44,3 |
La familia más numerosa en cuanto al número de géneros y de especies
es Asteraceae.
En la Tabla 1 se presentan las 14 familias representadas por cinco géneros o
más; el resto de familias están representadas por menos de cinco géneros de la
siguiente manera: siete familias con cuatro géneros, 10 familias con tres
géneros, 11 familias con dos géneros y 51 familias con un género (Figura 2).
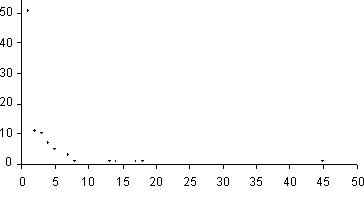 |
Figura 2. Número de géneros por familia en comparación con el número de familias.
En cuanto al número de especies, 22 familias están representadas por 20 especies o más (Tabla 2). La familia más rica en especies es sin duda Asteraceae con 251 especies, seguida por Melastomataceae y Solanaceae con más de 100 especies cada una, y Ericaceae, Rubiaceae y Campanulaceae con más de 50 especies. Del resto de familias es interesante anotar que 23 están representadas por una especie (Figura 3).
En cuanto a los géneros, 27 están representados por más de 15
especies (Tabla 3). El género más numeroso es
Miconia con 99 especies, seguido por
Piper con 43 y
Solanum con 40 especies.
Veinticuatro géneros están representados por 15 a 34 especies; del resto es
interesante anotar que 114 géneros están representados por dos a cinco especies
y 102 géneros lo hacen sólo con una especie (Fig. 4), incluidos 8 monotípicos
(Desfontainiaty, Gaiadendron, Margyricarpus, Pineda, Rhynchotheca, Schmardaea,
Streptsolen y Tessaria).
| Familia | Especies | % | Familia | Especies | % |
|---|---|---|---|---|---|
| Asteraceae | 251 | 16,0 | Scrophulariaceae | 29 | 1,8 |
| Melastomataceae | 170 | 10,8 | Araliaceae | 27 | 1,7 |
| Solanaceae | 106 | Cunoniaceae | 25 | 1,6 | |
| Ericaceae | 99 | 6,7 | Polygalaceae | 25 | 1,6 |
| Rubiaceae | 75 | 6,3 | Lauraceae | 25 | 1,6 |
| Campanulaceae | 56 | 4,8 | Boraginaceae | 24 | 1,5 |
| Piperaceae | 47 | 3,5 | Onagraceae | 22 | 1,4 |
| Piperaceae | 43 | 3,0 | Rosaceae | 22 | 1,4 |
| Euphorbiaceae | 36 | 2,7 | Myrsinaceae | 20 | 1,3 |
| Fabaceae | 36 | 2,3 | Symplocaceae | 20 | 1,3 |
| Clusiaceae | 34 | 2,2 | |||
| Berberidaceae | 32 | 2,0 | Otras familias | 344 | 21,9 |
Algunos géneros, si bien son nativos de zonas de vegetación más
bajas, frecuentemente se encuentran cultivados en los valles altos de la Sierra;
éstos son: Cyphomandra,
Nicotiana y
Schinus. Otros al parecer han sido
introducidos: Crataegus es un género
templado holártico que aparentemente fue introducido desde México en épocas
coloniales y Parajubaea cocoides
podría ser igualmente una planta cultivada.
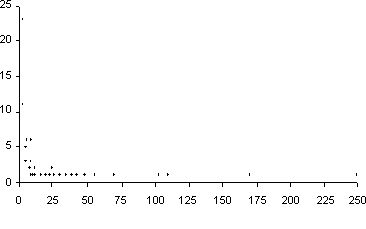 |
Figura 3. Número de especies por familia en comparación con el
número de familias.
| Género | Especies | % | Género | Especies | % |
|---|---|---|---|---|---|
| Miconia | 99 | 6,3 | Salvia | 21 | 1,3 |
| Piper | 43 | 2,7 | Lupinus | 20 | 1,3 |
| Solanum | 40 | 2,6 | Symplocos | 20 | 1,3 |
| Centropogon | 34 | 2,2 | Oreopanax | 19 | 1,2 |
| Baccharis | 32 | 2,0 | Meriania | 18 | 1,2 |
| Berberis | 32 | 2,0 | Hypericum | 17 | 1,1 |
| Calceolaria | 29 | 1,9 | Pentacalia | 17 | 1,1 |
| Gynoxys | 28 | 1,8 | Ageratina | 16 | 1,0 |
| Palicourea | 28 | 1,7 | Gaultheria | 16 | 1,0 |
| Monnina | 25 | 1,6 | Verbesina | 16 | 1,0 |
| Weinmannia | 25 | 1,6 | Cestrum | 15 | 1,0 |
| Fuchsia | 22 | 1,5 | Siphocampylus | 15 | 1,0 |
| Diplostephium | 21 | 1,3 | |||
| Brachyotum | 21 | 1,3 | Otros géneros | 879 | 56,0 |
De los datos aquí presentados, se puede decir que la mayoría de las familias están representadas por un solo género y así mismo numerosas familias están representadas por pocas especies, como se puede apreciar en las figuras 2 y 3. De igual manera, la mayoría de los géneros están representados por una o pocas especies, como se aprecia en la figura 4.
Si observamos el lado opuesto de los gráficos, tenemos que
una familia, Asteraceae, está representada por 44 géneros y 251 especies; seguida por
Melastomataceae, Solanaceae, Ericaceae y Rubiaceae con 13-17 géneros y 75-170 especies,
es decir que el 36% de los géneros (104) y el 45% de las especies (701) de árboles y
arbustos pertenecen a cinco familias. Melastomataceae, se ubica en quinto lugar en
cuanto al número de géneros (Tabla 1), pero en segundo lugar en cuanto al número de
especies (Tabla 2), que claramente está en relación con el alto número de especies
pertenecientes al género, Miconia (Tabla
3).
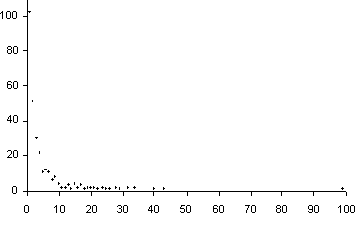 |
Figura 4. Número de especies por género en relación con el número de géneros.
Hay que anotar que los números de especies hasta hoy registrados, se basan en los conocimientos que disponemos al momento. Revisiones críticas de varios géneros grandes como Miconia, Berberis, Lupinus, Oreopanax, entre otros, seguramente alterarán estas cifras, pero al momento no podemos predecir en que proporciones.
Otros inventarios de regiones montañosas neotropicales
Se tomaron los resultados de las listas florísticas de dos regiones montañosas neotropicales, con vegetación similar a la del Ecuador, para hacer una comparación en cuanto a la diversidad.
Estas listas son de la flora leñosa de la zona alta (2000-3819 m) de la Cordillera de Talamanca, Costa Rica (Kappelle et al., 1991) y de las plantas de la zona alta (2300-4200 m) del Parque Nacional Río Abiseo, Perú (Young y León, 1990) (Figura 5.).
En Talamanca las familias más numerosas fueron Rubiaceae, Melastomataceae, Lauraceae, Asteraceae y Ericaceae, y los géneros más numerosos Chusquea, Miconia, y Ocotea. En Río Abiseo las familias más importantes numéricamente fueron Melastomataceae, Asteraceae, Solanaceae, Myrsinaceae y Ericaceae, y los géneros Miconia, Solanum y Schefflera.
Se puede decir que los taxa de leñosas numéricamente importantes en las tres áreas son Asteraceae, Melastomataceae, Ericaceae y Miconia. Miconia aparece desproporcionadamente numeroso en Río Abiseo (32 especies seguido por Solanum con 14) al igual que en el Ecuador, mientras en Costa Rica se ubica luego de Chusquea.
Patrones fitogeográficos de la flora leñosa andina
Humboldt (1807) estableció las bases de la distribución geográfica de las plantas en su "Essai sur la géographie des plantes", en base de sus observaciones en América tropical.
Estudios fitogeográficos posteriores sobre plantas andinas del Ecuador no se conocen hasta la publicación de Diels (1937). Al observar la vegetación del límite de los árboles, Diels anotó que lo que debería designarse florísticamente como elemento tropical asciende con la masa cerrada de la selva hasta niveles increíblemente altos. El autor se detuvo a analizar su denominado nivel andino alto que corresponde a la vegetación libre de árboles, y buscó los lugares de procedencia de algunos géneros para explicar que la flora altoandina no procede únicamente de la antigua flora tropical americana sino que procede de raíces muchos más amplias. Así, Diels designó algunos géneros como elementos boreal, antártico y cosmopolita según sus patrones de distribución. También discutió con algunos ejemplos, las relaciones de la flora altoandina ecuatorial con aquella de los Andes meridionales y de otras regiones montañosas australes y boreales.
Estudios fitogeográficos propiamente de los bosques andinos se conocen de la Cordillera Oriental (van der Hammen y Cleef 1983b) y de la Sierra Nevada de Santa Marta (Cleef et al., 1984, com. pers.) en Colombia. Ambos estudios proveen las bases para los estudios sobre el origen y la composición de la flora leñosa andina ecuatorial. El presente estudio es por lo tanto, la primera contribución al conocimiento fitogeográfico de la flora leñosa de los bosques andino-altoandinos del Ecuador.
Metodología. - El
análisis a nivel de géneros ha demostrado ser el más adecuado para los estudios
fitogeográficos y para este análisis se siguió la metodología propuesta por Cleef
(1979): para cada género se registró el número total de especies y el área de
distribución geográfica actual (estos datos se presentan en el octavo capítulo);
los géneros con áreas de distribución similares se agruparon en elementos
florísticos geográficos. Hay que recalcar que, los elementos fitogeográficos,
se basan en la distribución actual de los géneros y las áreas donde tiene la
mayor diversidad de especies se consideran, en la mayoría de los casos, como la
posible área de origen de donde un género colonizó los Andes tropicales. Si
bien para algunos géneros y especies se han hecho estudios filogenéticos, aún
no se dispone de datos suficientes para determinar elementos florísticos
genéticos (Cleef, 1979; Simpson y Todzia, 1990), más aún si todavía no se ha
superado la etapa de recolección de datos e identificación de las especies de
los Andes tropicales.
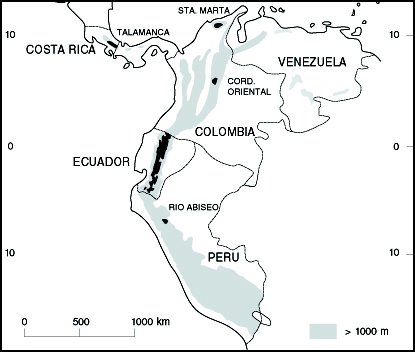 Figura 5. Mapa de ubicación de los sitios mencionados en el
texto, donde se han realizado inventarios florísticos y/o análisis
fitogeográficos.
Figura 5. Mapa de ubicación de los sitios mencionados en el
texto, donde se han realizado inventarios florísticos y/o análisis
fitogeográficos.
|
Elementos fitogeográficos
El presente análisis incluye 255 géneros de árboles y de arbustos andino-altoandinos y se reconocen nueve elementos fitogeográficos (Figura 6) que corresponden a tres componentes principales, de acuerdo con Cleef (1979) y van der Hammen y Cleef (1983a,b). Al componente tropical pertenecen los elementos andino tropical, neotropical, pantropical, americano-africano y americano-asiático; al componente templado pertenecen los elementos holártico, austral-antártico y ampliamente templado; y finalmente el componente cosmopolita está conformado por el elemento cosmopolita.
En el presente análisis no se ha considerado a los géneros del bosque subandino, que se encuentran en la zona de transición, tampoco aquellos propios de zonas secas o aparentemente cultivados -éstos géneros se mencionan en capítulos anteriores y están en el capítulo de las descripciones. Los resultados se comparan con los datos disponibles de los análisis de otras regiones montañosas neotropicales.
Elemento andino tropical.
- Incluye a los géneros de América tropical, cuya distribución principal se limita
a los Andes tropicales -Venezuela hasta el norte de la Argentina. Cuarenta y cinco
géneros, i.e. cerca del 18%,
pertenecen al elemento andino tropical. Los géneros incluidos son:
| Aequatorium | Critoniopis | Heppiella | Pineda |
| Aristeguietia | Cronquistianthus | Iochroma | Plutarchia |
| Asplundianthus | Cuatrecasanthus | Joosia | Polylepis |
| Badilloa | Delostoma | Joseanthus | Rhynchotheca |
| Brachyotum | Diogenesia | Killipia | Saracha |
| Brugmansia | Dioicodendron | Kingianthus | Schmardaea |
| Bucquetia | Dunalia | Llerasia | Semiramisia |
| Cacosmia | Espeletia | Loricaria | Steiractinia |
| Cantua | Ferreyranthus | Margyricarpus | Streptosolen |
| Ceroxylon | Geissanthus | Minthostachys | Vallea |
| Cervantesia | Grosvenoria | Monactis | |
| Columellia | Gynoxys | Pappobolus |
La mayor parte de este elemento lo representa la familia Asteraceae con 19 géneros. Dos géneros incluidos aquí son propios del páramo y ocasionalmente pueden estar entremezclados con elementos del bosque altoandino: Espeletia es un elemento paramuno (Cleef, 1979) y Loricaria es propio del páramo y la puna. Bucquetia es característico del (sub)páramo de Venezuela, Colombia y Ecuador pero también se encuentra dentro del bosque. Un solo género, Kingianthus, es endémico del Ecuador. Joseanthus (Vernonia, Killipia y Plutarchia están representados sólo en Colombia y Ecuador, éste último fue considerado un elemento paramuno por Cleef (1979), pero en el Ecuador no es exclusivamente un género del páramo. Taxa representados en el centro-sur del Ecuador y norte-centro del Perú son: Cuatrecasanthus (Vernonia), Ferreyranthus, Grosvenoria, Monactis, Pineda, Rhynchotheca y Streptosolen. Los dos géneros de Rosáceas, Margyricarpus y Polylepis, también se encuentran en la región templada austral; ambos pertenecen a la tribu Sanguisorbeae, mejor desarrollada en el hemisferio austral (Simpson, 1986).
Elemento neotropical.
- En este elemento se incluyen los géneros que también se distribuyen en otras áreas
de América tropical o subtropical. Dos grupos pueden distinguirse: a) aquellos que
son más diversos en las zonas montañosas y b) aquellos que son más diversos en las
tierra bajas. Ciento trece géneros pertenecen al elemento neotropical, i.e.
cerca del 44%, de los que noventa son géneros montanos (35%) y veintitrés son de las
tierras bajas (9%). Entre los géneros montano-neotropicales se encuentran los
siguientes:
| Abatia | Chusquea | Koanophyllon | Phenax |
| Aegiphila | Cinchona | Ladenbergia | Psammisia |
| Ageratina | Clusia | Lepechinia | Psidium |
| Aiphanes | Coutarea | Liabum | Purdiaea |
| Aphelandra | Cuatresia | Llagunoa | Rhipidocladum |
| Arcytophyllum | Cyathea | Lozanella | Roupala |
| Aulonemia | Cybianthus | Macleania | Ruagea |
| Axinaea | Dalea | Macrocarpaea | Salpichroa |
| Ayapanopsis | Dasyphyllum | Mauria | Schistocarpha |
| Baccharis | Deprea | Meriania | Sessea |
| Barnadesia | Diplostephium | Miconia | Siparuna |
| Bejaria | Disterigma | Monnina | Siphocampylus |
| Bocconia | Duranta | Monochaetum | Smallanthus |
| Brunellia | Elaeagia | Myrcianthes | Sphyrospermum |
| Burmeistera | Freziera | Nectandra | Styloceras |
| Calea | Gaiadendron | Nephelea | Symbolanthus |
| Capsicum | Gaylussacia | Oligactis | Tecoma |
| Carica | Graffenrieda | Ophryosporus | Themistoclesia |
| Cavendishia | Hesperomeles | Oreopanax | Thibaudia |
| Cedrela | Hypericum | Orthaea | Tibouchina |
| Centropogon | secc. Brathys | Palicourea | Verbesina |
| Ceratostema | Huberia | Panopsis | |
| Cestrum | Kaunia | Pentacalia | |
| Critoniopsis |
Arcytophyllum, Diplostephium, Gaiadendronz, Hesperomeles, Cinchona y Mauria prácticamente tienen una distribución andino-tropical ya que sólo una o dos especies se encuentran en las montañas centroamericanas y probablemente se trasladaron desde los Andes (Figura 7). Baccharis y Verbesina si bien están centrados en las montañas neotropicales, se encuentran ampliamente distribuidos en toda América. Hypericum tiene una distribución actual ampliamente templada, pero la sección Brathys -sección americana donde se encuentran todas las especies leñosas representadas en los Andes del Ecuador- se habría ramificado tempranamente del tronco original en el oeste de Gondwana (Africa) y posteriormente se diversificó en América tropical (Robson, 1987). Dicha sección tiene dos centros primarios: el primero en Cuba y Belice, y el segundo en la frontera colombo-venezolana, de ésta última área se diversificó la porción arbustiva hacia el norte y por los Andes hasta Bolivia (Robson, 1990).
Un par de géneros que tiene su mayor diversidad en otras áreas de América tropical, están representados con una especie en la región andina. Este es el caso de Huberia con nueve especies en el sureste del Brasil y sólo H. peruviana en los Andes del sur del Ecuador y norte del Perú; y Purdiaea con once especies en Cuba y sólo P. nutans en los Andes del norte.
En los análisis de Colombia y Costa Rica, Miconia y Bocconia han sido listados dentro del elemento americano-africano y americano-asiático, respectivamente y removidos al elemento montano neotropical en este estudio. La única especie africana de Miconia, aparentemente fue descrita en base a material brasilero erróneamente etiquetado (S. Renner, com. pers.; Wurdack, 1970) y Bocconia es un género netamente neotropical (M. Lidén, com. pers.).
Veintitrés géneros ampliamente distribuidos en las tierras bajas de
América tropical penetran en el bosque andino y en ocasiones una o pocas especies
pueden encontrarse hasta los 3000 metros de altitud. Estos géneros son:
| Aiouea | Citharexylum | Geonoma | Lepidaploa |
| Alloplectus | Clibadium | Gonzalagunia | Myrcia |
| Austroeupatorium | Coursetia | Hoffmannia | Ossaea |
| Besleria | Daphnopsis | Hyeronima | Trichipteris |
| Blakea | Faramea | Inga | Zinowiewia |
| Chamaedorea | Gasteranthus | Leandra |
Aiouea, género característico de la cuenca Amazónica, está representado por una especie propia de los bosques andinos: Aiouea dubia. Géneros característicos, pero no exclusivos de los bosques andinos son: Cedrela, Clusia, Hyeronima, Miconia, Myrcianthes, Roupala, Siparuna y Styloceras. En el caso de Miconia, es interesante el hecho de que es el género con mayor número de especies (99) registradas en este estudio. Cuatresia recientemente ha sido recolectado en los bosques andinos, lo que amplía su distribución altitudinal sobre los bosques subandinos y Zinowiewia últimamente se ha registrado en el sur del Ecuador.
Elemento pantropical.
- En esta categoría se encuentran los géneros ampliamente distribuidos en América
tropical y en los trópicos de Africa y Asia. Treinta géneros, i.e. cerca del 12%, tienen una
distribución pantropical:
| Acalypha | Coreopsis | Mimosa | Schefflera |
| Alchornea | Croton | Myrsine | Senna |
| Allophylus | Desmodium | Otholobium | Ternstroemia |
| Buddleja | Dodonaea | Pilea | Tournefortia |
| Byttneria | Eugenia | Phyllanthus | Zanthoxylum |
| Casearia | Heliotropium | Piper | |
| Cleome | Justicia | Psychotria | |
| Cordia | Maytenus | Sapium | |
| Ficus |
Varios de estos géneros están ampliamente distribuidos desde las tierras bajas hasta los bosques andinos. Casearia un género propio de las tierras bajas, está representado por una especie común en los bosques andinos del norte del país: Casearia mexiae. Otros como Buddleja son muy diversos en los Andes y pueden encontrarse hasta por sobre los 4000 metros de altitud, y sólo pocas especies descienden bajo los 2000 metros; Otholobium (Psoralea) tiene, en Sur América tropical, una distribución andina y generalmente se encuentra sobre los 2000 metros de altitud (Grimes, 1990); Mimosa ser. Andinae se extiende en la región interandina desde el sur de Colombia hasta el norte del Perú (Barneby, 1991).
Elemento americano-africano. - Tres géneros, i.e. cerca del 1%, tienen su distribución en los trópicos de América y Africa. Estos géneros se denominan también tropicales amfi-atlánticos o afro-americanos, y son:
| Guarea | Lantana | Ocotea |
Los tres géneros tienen numerosas especies en las tierras bajas de América tropical y pocas en Africa. En los bosques andinos están representados por pocas especies.
Elemento americano-asiático.
- En este grupo se encuentran los géneros que en la actualidad se distribuyen
principalmente en los trópicos de América y el sureste de Asia y Malasia.
Estos géneros se denominan también tropicales amfi-pacíficos o malayo-americanos.
Diecisiete géneros, i.e.
cerca del 7%, pertenecen a este elemento:
| Citronella | Hedyosmum | Saurauia | Turpinia |
| Clethra | Lycianthes | Schoepfia | Xylosma |
| Dicksonia | Meliosma | Sphaeropteris | |
| Gordonia | Persea | Styrax | |
| Guettarda | Pouteria | Symplocos |
Styrax está representado en la región mediterránea; Clethra y Persea se encuentran también en el sureste de los Estados Unidos y tiene una especie en Madeira y las Islas Canarias respectivamente. Gordonia y Meliosma tienen especies en la región templada del norte y Dicksonia en Australia, Nueva Zelanda y Juan Fernández. Hedyosmum tiene una sola especie en el sureste de Asia y el resto en América tropical, especialmente en los Andes. Pouteria está pobremente representado en los bosques andinos.
Elemento holártico. - En
este elemento se incluyen los géneros cuya distribución principal se encuentra en el
hemisferio boreal templado y se extienden por las zonas frías de las montañas
tropicales hacia el hemisferio austral. Veintidós géneros pertenecen al
elemento holártico, i.e. cerca del
9%. Géneros incluidos en este elemento son:
| Alnus | Berberis | Juglans | Myrica |
| Ambrosia | Boehmeria | Lupinus | Prunus |
| Astragalus | Cornus | Morus | Rhamnus |
| Ribes | Salvia | Scutellaria | Viburnum |
| Rubus | Sambucus | Vaccinium | |
| Salix | Satureja | Valeriana |
Scutellaria es un género subcosmopolita pero la sección Scutellaria tiene su mayor diversidad en las montañas de México y los Andes del norte. En los Andes dicha sección se habría diversificado en los últimos 5-3 millones de años luego de ingresar por el istmo de Panamá y posteriormente lo habría hecho la sección andina Perilomia, durante el pleistoceno (Paton, 1990). Igualmente Valeriana habría migrado desde Norteamérica hacia los Andes, donde tiene su actual centro de diversidad, luego de la formación del puente de tierra entre Norte y Sur América (Larsen, 1986; Eriksen, 1989). Astragalus se encuentra en Sur América desde el Ecuador hasta el extremo sur del continente; al parecer las especies sudamericanas son el producto de una antigua migración hacia el sur a partir de un ancestro originario de Norte América (Johnston, 1947) (Figura 7).
Elemento austral-antártico.
- A este elemento corresponden los
géneros que tiene su distribución principal en las regiones frías y moderadamente
frías del hemisferio austral e incluye América del Sur templada e islas adyacentes;
la región subantártica y antártica; Tasmania, Nueva Zelanda y Australia;
sureste de Asia y ocasionalmente extendiéndose hacia el norte hasta China y
Japón (Cleef, 1979). En América tropical estos géneros se extienden por las
regiones montañosas. Diecinueve géneros, i.e.
cerca del 7%, pertenecen a este elemento. De éstos, tres géneros están ampliamente
distribuidos en la región andino-austral-antártica y son:
| Acaena | Podocarpus | Weinmannia |
Siete géneros están distribuidos en América tropical y en Austral-Asia:
| Drimys | Gaultheria | Oreocallis | Prumnopitys |
| Fuchsia | Lomatia | Pernettya |
Y, finalmente nueve géneros están distribuidos en América tropical
y Suramérica templada. Estos géneros americanos son:
| Blepharocalyx | Colletia | Myrteola |
| Calceolaria | Desfontainia | Tristerix |
| Chuquiraga | Escallonia | Ugni |
Chuquiraga, Myrteola y Ugni son característicos de la vegetación achaparrada del subpáramo; los dos primeros se extienden hacia los páramos. Blepharocalyx y Ugni son dos géneros bastante raros en el sur del Ecuador. Desfontainia es un género monotípico que se extiende desde Costa Rica hasta el estrecho de Magallanes. Colletia, Lomatia y Oreocallis tienen su distribución más septentrional en los Andes del Ecuador, mientras que Chuquiraga llega hasta el sur de Colombia. La ubicación de Drimys en el elemento austral-antártico podría trasladarse con futuros estudios: antepasados de las Winteraceae al parecer tuvieron una distribución tropical más amplia en el pasado y la distribución actual austral templada la habrían alcanzado posteriormente (Doyle et al., 1990).
Elemento ampliamente templado. - En este elemento se incluye a los géneros ampliamente distribuidos en las regiones templadas de ambos hemisferios y en las zonas frías de las montañas tropicales. Dos géneros Coriaria y Ephedra (i.e. cerca del 1%) pertenecen al elemento ampliamente templado. Ephedra fue listado por Gentry (1982) con inseguridad, como un género laurásico y Coriaria como templado del sur. Este último género presenta una distribución tan fragmentada que difícilmente puede asignarse a uno de los elementos anteriores.
Elemento cosmopolita. - Cuatro géneros, i.e. el 1%, tienen una distribución prácticamente cosmopolita, o subcosmopolita. Estos géneros están bien representados en las tierras bajas y montañas tropicales y en las regiones templadas. Entre éstos se encuentran Blechnum, Euphorbia, Ilex y Solanum. Euphorbia laurifolia es la única especie arbórea de ese género registrada en este estudio. Blechnum está representado por dos especies arborescentes, mientras que Solanum es uno de los géneros más diversos, representado con 40 especies leñosas en los Andes del Ecuador.
Comparación con otras regiones montañosas neotropicales
Los resultados aquí obtenidos se compararon con los datos fitogeográficos disponibles de tres regiones montañosas neotropicales con vegetación similar. Estas áreas corresponden a los bosques montanos de roble, Quercus, (2000-3400 m) de la Cordillera de Talamanca en Costa Rica (Kappelle et al., 1992) y, a los bosques andinos ( ca. 2300-3500 m) de la Cordillera Oriental (van der Hammen y Cleef, 1983b) y la selva andina-bosques andinos (2500-3300 m) de la Sierra Nevada de Santa Marta, ambos en Colombia (Cleef et al., 1984; Cleef, com. pers.) (Figura 5). El número de géneros registrados en Talamanca (166), Santa Marta (64,), Cordillera Oriental (130) y el Ecuador (255) es muy variable en gran parte debido a los distintos métodos de muestreo, a las áreas consideradas y al propósito de los estudios, además los muestreos de Colombia son bastante incompletos (Cleef, com. pers.), por lo tanto la comparación que aquí se presenta es preliminar.
Al comparar los espectros fitogeográficos de las cuatro áreas se puede apreciar que son bastante similares (Figura 6, Tabla 4).
El componente tropical (elementos neotropical incluido el andino, pantropical, americano-asiático y americano-africano) ocupa un alto porcentaje: el 81% en Talamanca, el 75% en Santa Marta, el 82% en la Cordillera Oriental y el 82% en el Ecuador.
Al analizar los porcentajes de cada elemento en detalle se puede apreciar que el elemento neotropical (incluido el andino) está mejor representado en el Ecuador, con un 10% más que en las otras áreas; sin embargo el elemento andino tropical tiene porcentajes similares en el Ecuador y en la Cordillera Oriental colombiana. En los cuatro estudios el elemento americano-asiático (amfi-pacífico o malayo-americano) es importante, pero es ligeramente más alto en Costa Rica que en los bosques andinos (más del 10% en Centroamérica y hasta el 9% en los Andes). Estos géneros, así como los elementos holárticos, habrían ingresado hacia los Andes desde el norte a través del puente de tierra de Centro América (van der Hammen y Cleef, 1983 a,b) (Figura 7), lo que explica el mayor porcentaje en Costa Rica.
Si se analiza el componente templado (elementos holártico,
austral-antártico y ampliamente templado), se puede apreciar que el elemento
holártico tiene un porcentaje más alto en los Andes que en Costa Rica, donde
se esperaría sea mayor, y posiblemente este resultado se deba a las diferencias
en los muestreos. El elemento austral-antártico tiene porcentajes similares en
Costa Rica y en los bosques andinos y al parecer los géneros incluidos en este
elemento pudieron llegar a las montañas de América Central utilizando los Andes
como corredor (Figura 7).

|
El porcentaje del elemento austral es ligeramente mayor en Costa Rica, sin embargo, en números absolutos, se encuentran más géneros austral-antárticos en el Ecuador que en Talamanca (19 versus 12) y la diferencia incluye géneros que alcanzan su distribución más septentrional en el Ecuador.
| ELEMENTOS |
ECUADOR N=255 |
C. ORIENTAL N=130† |
STA. MARTA N=64† |
Talamanca N=166 |
|---|---|---|---|---|
Holártico |
9 |
10 |
14 |
7,8 |
| Austral-antártico | 7 | 6 | 8 | 7,2 |
| Ampliamente templado | 1 | 1* | - | 2,4 |
| Cosmopolita | 1 | 1* | 3 | 1,8 |
| Americano-asiático | 7 | 8 | 9 | 10,8 |
| Americano-africano | 1 | 4 | - | 4,2 |
| Pantropical | 12 | 19 | 19 | 12,7 |
| Neotropical | 44 | 34 | 33 | 53 |
| Andino tropical | 18 | 17 | 14 | - |
| Total | 100 | 100 | >100 | 99,9 |
†inventarios incompletos (Cleef, com.pers.);*aproximado basado en Cleef et al. (1984, Figura 88: 331).
La gran similitud de la flora montana de Costa Rica con la de los Andes colombianos ya fue estudiada por Cleef y Chaverri (1992) y Kappelle et al. (1992). Si bien los bosques de Talamanca están dominados por robles del género Quercus, ausente en el Ecuador, es interesante anotar el alto porcentaje de géneros compartidos en ambas zonas.
De los 166 géneros de leñosas de Talamanca, más del 60% se comparte con el Ecuador -en el subcapítulo anterior también se recalcó que las familias de leñosas más numerosas son compartidas entre el bosque de roble de Costa Rica y los bosques andinos del Ecuador. En el porcentaje de géneros no compartidos se encuentran varios ausentes en Sur América, como son, por ejemplo, Cleyera (Theaceae), Comarostaphylis (Ericaceae), Garrya (Garryaceae), Hemichaena (Scrophulariaceae), Lopezia (Onagraceae), Mahonia (Berberidaceae) y y Microtropis (Celastraceae). También se incluyen aquellos que se encuentran en los bosques montanos y/o páramos de Costa Rica y Colombia -en este caso en Talamanca y Santa Marta- y no se distribuyen hacia el sur del continente como, por ejemplo, Chaetolepis (Melastomataceae) y Holodiscus (Rosaceae). Alfaroa (Juglandaceae), Matudaea (Hamamelidaceae) y Quercus (Fagaceae) si bien entran desde América Central hasta Colombia, no se encuentran en Santa Marta.
La mayoría de los 64 géneros de leñosas de los bosques andinos de la Sierra Nevada de Santa Marta es compartida con el Ecuador (54 géneros o el 86%). Este porcentaje incluye solamente dos géneros andino-tropicales (Ceroxylon y Vallea), es decir que el resto pertenecen a los otros elementos.
En el porcentaje no compartido se encuentran, además de los antes mencionados en común con Talamanca, Killipiodendron (Theaceae) endémico de Colombia, Paragynoxys (Senecioneae, Asteraceae) y Libanothamnus (Espeletiinae, Asteraceae), género colombo-venezolano de los (sub)páramos. En general, las Espeletiinae tienen su mayor diversidad en los Andes de Colombia y Venezuela ya que de las 130 especies sólo Espeletia pycnophylla, ingresa al Ecuador (Cuatrecasas, 1986).
En general se puede decir que el elemento principal de los bosques
alto montanos en los Andes del Norte y Costa Rica, es el neotropical, seguido por el
pantropical, el holártico y el austral-antártico (Figura 6). El elemento
americano-asiático es importante, mientras que el elemento americano-africano
apenas sobrepasa el 4%, y el cosmopolita escasamente llega al 3%.

|
La vegetación del sur del Ecuador presenta una flora bastante diversa y aún poco conocida en mucha áreas. Varios géneros distribuidos en los Andes y en otras regiones montañosas de Centro y Sur América, se conocen en el Ecuador solamente en el sur. Por ejemplo se pueden mencionar los siguientes géneros: Bejaria, Blepharocalyx, Bucquetia, Dioicodendron, Drimys, Purdiaea, Schoepfia y Ugni. Son por lo tanto necesarios estudios de los eventos geológicos, tipo de suelo, hábitat, polinización, dispersión, etc., para tratar de explicar este interesante patrón de distribución.
En resumen, la flora leñosa de los bosques andinos y altoandinos del Ecuador está conformada por un poco más del 50% de géneros neotropicales centrados en los Andes: el 18% de géneros netamente andinos tropicales y el 35% de los géneros del elemento montano-neotropical. Los Andes ecuatorianos son también el límite septentrional de varios géneros representados hacia el sur del continente. El futuro análisis de la flora vascular en general proporcionará mayores detalles sobre los patrones de migración de la riquísima flora de los bosques andino-altoandinos de los Andes del Ecuador.