J. ARNOLD ARBOR. 61: 41-94. 1980.
Shiu Ying Hu
MY FIRST ASSOCIATION with Metasequoia glyptostroboides Hu & Cheng was at the Arnold Arboretum of Harvard University during the winter of 1948-1949. At that time help was needed to place fresh, recently imported metasequoia seed into small envelopes for distribution to botanical institutions, forest experiment stations, and interested individuals around the world. These seeds, mailed from Nanking, China, on November 29, 1948, and totaling 500 grams, were the second shipment received at the Arnold Arboretum. A letter from Professor W. C. Cheng concerning the shipment was later placed with the unmounted specimens of M. glyptostroboides that had been collected during the summer of 1948.
In addition to the seed, five shipments of herbarium specimens collected in southwestern Hupeh Province between 1946 and 1948 were received at the Arnold Arboretum by the late Professor Elmer D. Merrill. In 1973 this herbarium material was turned over to me for identification, and this article is concerned with the information resulting from that undertaking. A systematic enumeration of all identifiable species represented in the five collections (an assemblage of species here termed the "metasequoia flora") is presented below. While the type locality of Metasequoia glyptostroboides is in eastern Szechwan, the specimens on which this work is based are from the general area (here referred to as the "metasequoia area") in Hupeh Province where M. glyptostroboides was later discovered growing in a 'natural population (see below). Also included are brief summaries of the expeditions that obtained the five collections, as well as a short account of earlier botanical collections from Central China, a description of the salient features of the metasequoia flora, an analysis of the gymnosperms that occur with metasequoia, and my interpretation of the metasequoia flora.
In 1950 K. L. Chu and W. S. Cooper published the results of an ecological reconnaissance of the metasequoia community. A posthumously published paper by E. H. Fulling (1976), along with additions published in 1977, summarizes the history of the discovery of Metasequoia glyptostroboides and presents an annotated bibliography of published references to metasequoia. Only information not available in these three readily available articles is included here.
In the enumeration of species, the system followed in the arrangement of the families conforms with that used in Iconographia Cormophytorurn Sinicorum (Anonymous, 1972-1976). For economy of space, and unless
(c) President and Fellows of Harvard College, 1980.
Journal of the Arnold Arboretum 61: 41-94. January, 1980.
[p. 41]
nomenclatural change is involved, only the name, important synonyms, and voucher specimens are cited. Assistance of specialists is recognized in parentheses after either the group or the species.
Earlier authors have employed different spellings for the same collectors or localities. For those who lack a knowledge of the geography of the region and the Chinese language, these differences are often confusing. In this article, the following guidelines have been used in deciding between alternative spellings of Chinese personal and place names. For collectors the spellings on the herbarium labels have been used. However, with regard to the name of the man who discovered metasequoia, earlier authors have used either T. Wang or C. Wang. Since his publication with P. Y. Fu on Salix (1974) appeared under his name as Zhan Wang, the initial of his given name adopted here is Z.
Concerning localities, for names of well-known places such as Chungking, Hupeh, and Szechwan, the widely recognized spellings of the Chinese Post Office and foreign presses have been adopted, and the Rand McNally World Atlas (Anonymous, 1949) has been used as a reference. Names of minor localities have been transliterated using the system of romanization outlined in the Harvard University Press edition of Mathew's (1931) A Chinese-English Dictionary. Cross-references to different spellings of various names that have already appeared in publications are given below,[1] where recognized spellings are given in boldface type. The names listed under "other localities" are alphabetically arranged.
BOTANICAL INVESTIGATIONS IN CENTRAL CHINA AND THE
METASEQUOIA AREA
The botanical expeditions that have contributed to our knowledge of the metasequoia flora are numerous. Some were mounted specifically for the investigation of the area in which Metasequoia glyptostroboides was discovered as an extant plant, while others were conducted for the general botanical exploration of the flora of Central China. Metasequoia glyptostroboides is now known to occur naturally in western Hupeh and adjacent Szechwan provinces southward to the hills of western Hunan (cf. Cheng, Fu, et al.,
[1]Type locality of Metasequoia glyptostroboides: Mo-tao-chi, Mo-tao-hsi (Hu & Cheng), Mou-tao-chi (Merrill, Hu).
Collector of syntypes: C. Y. Hsüeh, C. J. Hsüeh (Hu & Cheng, Merrill, Florin), C. Y. Hsieh (Hu).
Metasequoia grove: Shui-sa-pa, Shui-hsa-pa (Chaney, Chu & Cooper, Florin).
Other localities: Chien-shih, Kien-shi (Hu & Cheng, map); Chung Hsien, Chunghsien (Merrill, map); Chungking, Chunking (Hu); En-shih, An-shi (Chu & Cooper, map), An-shih (Hu & Cheng, map); Feng-chieh, Fungkien (Merrill, map); Hsiao-ho, Hsio-ho (Merrill, map), Shia-ho (Chu & Cooper, map); Hung-pan-ying, Houng-pin-ying (Merrill), Huong-pan-yin (Chu & Cooper); K'ai Hsien, Kaisien (Merrill, map); Li-chuan, Li-chuanhsien (Hu & Cheng, Merrill), Li-chwan (Hu & Cheng, map); Wan Hsien, Wanhsien (Hu & Cheng), Wan-hsien (Merrill); Wang-chia-ying, Wang-jia-ying (Hwa 182, field note), Wang-kia-ying (Hu & Cheng, Merrill); Yangtze River, Yang Tze Kiang (Hu & Cheng, map); Yun-yang, Yuyang (Merrill, map).
[p. 42]
1978, p. 310). It is of interest to note that several well-known collectors came within kilometers of the metasequoia area (MAP 1), but none penetrated it until 1943, the year when Z. Wang (FIGURE 1) first collected Metasequoia.
EARLIER COLLECTIONS FROM CENTRAL CHINA. Many historical collecting localities are within a 250 kilometer radius of the metasequoia area (MAP 1) and our present knowledge of the flora of Central China is largely the result of the expeditions and collecting activities of both western and Chinese botanists within this larger area. Europeans and Americans who had collected in Central China before the discovery of Metasequoia glyptostroboides include Thomas Watters, Augustine Henry, E. H. Wilson, P. Farges, A. von Rosthorn, A. C. Steward, the French missionaries E. Bodinier, J. Cavalerie, and J. Esquirol, and the Italian missionary C. Silvestri.
The many species new to science that were included in the collections made by these botanists were, likewise, described by both American and European taxonomists. Notable among this group were H. F. Hance, W.
[p. 43]
B. Hemsley, D. Oliver, A. Rehder, E. Koehne, C. Schneider, A. Franchet, H. Léveillé, and R. Pampanini.
Active floristic investigations carried out by Chinese botanists in Central China did not begin until western-trained students of botany returned to China and became eminent in that country's educational institutions. Several of the Chinese botanists who received their training at the Arnold Arboretum, and whose activities have advanced our knowledge of the metasequoia flora either directly or indirectly, are mentioned here. The first was Professor S. S. Chien, who, immediately after his return to China, taught in the National Southeastern University of Nanking (later renamed National Central University and now merged with Nanking University). In the late 1920's he was responsible for developing the botanical collections of the newly established Biological Laboratories of the Science Society of China. One of his goals was to establish a reference herbarium; toward this end, he sent one of his students, Y. L. Keng, to collect in Kiangsu and Chekiang in 1927. In the spring of 1928, he sent W. P. Fang to Szechwan to collect at Chin-fu-shan and many other localities (MAP 1). Fang was born and raised in Chung Hsien, 60 kilometers from Mo-tao-chi, and only 75 kilometers west of the metasequoia area. Although he stopped and collected in Chung Hsien, he did not reach the metasequoia area. Duplicates of Fang's collections are deposited in the herbarium of the Arnold Arboretum.
H. H. Chung was another Arnold Arboretum-trained botanist, who, in the early 1930's, became Professor of Botany in the National Wu-Han University. In 1934 he sent Ho-chang Chow (= H. C. Cheo, who had accompanied A. N. Steward on expeditions to Kweichow and Kwangsi in 1931 and 1933, respectively) to botanize southwestern Hupeh Province. Chow (MAP 1) began to collect in Pa-tung in early May, and, to judge from data on his specimens, he went on to collect in Chien-shih in August. By September he had reached En-shih, a locality only 60 kilometers east of the metasequoia area. A set of Chow's collections is also in the Arnold Arboretum herbarium.
Professor H. H. Hu, the botanist who identified Z. Wang's collection as belonging to the genus Metasequoia Miki and who became senior author of the name Metasequoia glyptostroboides, was a pioneer Chinese student at the Arnold Arboretum and the first Chinese botanist to receive a doctorate from Harvard University. His loyalty to the Arnold Arboretum and his continuous collaboration with E. D. Merrill contributed to the discovery and subsequent wide distribution of metasequoia.
The changes brought about by the Sino-Japanese War (1937-1945) had a definite effect leading to the discovery of Metasequoia glyptostroboides. The geographic isolation and the lawlessness associated with the border area where M. glyptostroboides is confined had reduced the population and helped to delay botanical exploration of the region. However, to improve wartime communications between the provincial capital of Hupeh at En-shih and the national capital at Chungking, a motor road was constructed in early 1939. Consequently, the formerly isolated hillside villages of Mo-tao-chi and Wang-chia-ying became rest stations, and the once inaccessible type locality of M. glyptostroboides and the surrounding region became more easily accessible.
[p. 44]
COLLECTIONS FROM THE METASEQUOIA AREA. The region from which the five collections discussed in this paper were gathered includes an area of approximately 800 square kilometers (cf. maps in Hu & Cheng, 1948; Chu & Cooper, 1950; and Merrill, 1948). Three-fourths of the specimens, however, are known to have been collected in the immediate vicinity of the metasequoia grove at Shui-sa-pa ("Water Fir Grove"); it is at this site that C. T. Hwa discovered a natural stand of mature, reproducing trees of Metasequoia glyptostroboides in the fall of 1947 (see below). According to Chu and Cooper, the metasequoia grove at Shui-sa-pa is located on a "strip [of land] along the main river 25 km. long and less than 1.5 km. wide. The altitude here ranges from 1,000 to 1,100 m." With the exception of Hwa's 1947 trip, so far as I am aware, all other expeditions to the area went straight to Shui-sa-pa.
The first shipment of specimens received at the Arnold Arboretum from the metasequoia area contained only two numbers: C. Y. Hsüeh 5, February 20, 1946, which consists of leafless branchlets with microsporangiate and mature megasporangiate cones of Metasequoia glyptostroboides, and Hsüeh 51, May 18, 1946, consisting of leafy shoots and immature strobili. These collections later became the syntypes of the species name. Z. Wang's collection of M. glyptostroboides, made in 1943, is deposited in the Herbarium, Bureau of Forestry Research, Ministry of Agriculture and Forestry, Government of China, and is not known in herbaria outside China.
The second shipment contained 179 specimens collected by C. T. Hwa during September, October, and November of 1947. Hwa's trip was financed by the Arnold Arboretum especially for the collection of metasequoia seed (Merrill, 1948). However, due to scant seed production, only a small amount was obtained. This was sent to the Arnold Arboretum, arriving there on January 5, 1948. The outstanding contribution of this expedition was neither the amount of seed nor the number of herbarium specimens collected, but the discovery of the location of an actively reproducing population of metasequoia at Shui-sa-pa in Hupeh Province.
During this trip Hwa gathered information from the local people about the occurrence of metasequoia, and he visited all known localities. In Wang-chia-ying, a large village in the Li-chuan District, he saw the largest known metasequoia tree, which measured about 50 meters in height, 3.3 meters in diameter across the basal buttresses, and 2.2 meters in diameter at breast height. Hwa's experiences and field observations laid the foundation for later explorers, and he served as guide for two of the three expeditions made during the spring and summer of 1948.
The third collection of specimens from the metasequoia area was made by Hwa in late February and early March, 1948, when he acted as guide for the expedition of Professor R. W. Chaney, paleobotanist and paleoecologist from the University of California, Berkeley, and Dr. M. Silverman, science writer for the San Francisco Chronicle. Although the party was in the metasequoia area for only five days, Hwa collected about 250 specimens. These were received at the Arnold Arboretum without labels, but each had a small tag bearing a number and Hwa's name in Chinese characters.
The fourth collection was made during the summer of 1948, when a team
[p. 46]
of Chinese specialists was organized by Professor W. C. Cheng for the intensive investigation of the flora and ecology of Shui-sa-pa. This expedition was financed by the American Philosophical Society under the joint sponsorship of Merrill and Chaney. Dr. Cheng was the leader of the group, Professor K. L. Chu was the ecologist, and Hwa once more served as guide. The specimens resulting from this expedition bear tags stamped "Hupeh-Szechwan, 1948, W. C. Cheng--C. T. Hwa," and the numerical sequence ranges from 550 to 1183. Three sets in ten bundles representing the results of the fourth expedition were dispatched, together with Hwa's earlier 1948 collections, to the Arnold Arboretum from Nanking by surface mail on December 29, 1948. The three sets do not have equal numbers of specimens. The Arnold Arboretum set contains 840 specimens that can be identified to species, although some of them include neither flowers nor fruits; in addition, there are 35 sterile specimens, which cannot be placed in genus or family. It would appear that some numbers were assigned in the laboratory after the specimens were sorted. All ferns have relatively high numbers, and specimens of closely related species are essentially in numerical sequence. After identification, duplicate sets were mailed to the Royal Botanic Gardens, Kew, and to the Herbarium of the University of California, Berkeley, in accordance with the arrangements made by Merrill and Cheng.
The fifth collection was made by Dr. J. L. Gressitt, of the Lingnan Natural History Museum and the California Academy of Sciences, who went with his assistant, Mr. Y. W. Djou, to conduct an entomological survey of the metasequoia area in late July, 1948. Gressitt left Shui-sa-pa in late August, while Djou remained there for an additional month. Since their specimens were collected to document the host plants of various insects, many have neither flowers nor fruits. A portion of the collection bears Gressitt's numbers 2401 to 2575, while the remainder have Djou's numbers 101 to 139.
In general, the quality of the specimens resulting from the five collecting trips described above is very poor. Most of the Hwa, Djou, and Gressitt numbers are sterile. Some of the Cheng-Hwa collections are sterile, while others are very inadequate, consisting in some instances of a single small herbaceous plant. Normally such specimens are not retained in research herbaria, but since our objective with these collections has been to document the flora of the metasequoia area as completely as possible, all specimens that could be determined to species have been retained and are included in the enumeration below. The majority of specimens have been named by comparison with types, isotypes, or photographs or fragments of type material in the herbaria of the Arnold Arboretum and Gray Herbarium, and a total of 550 species in 301 genera representing 127 families have been recognized in the five combined collections from the metasequoia area.
Despite the documentation of the metasequoia flora thus made possible, a complete flora of the area has not been forthcoming. Chaney (1948) reported that small fan palms (Trachycarpus fortunei (Hooker f.) H. Wendl.) grow at various places in Shui-sa-pa, while Chu (in Chu & Cooper, 1950, p. 273) observed that in a quadrat placed on muddy seepage ground, the surface was covered by a soft carpet of Diarrhena sp. (Gramineae). Neither taxon,
[p. 47]
however, is represented in the Arnold Arboretum material. In general, foresters, dendrologists, and entomologists pay little attention to the grasses and palms, which may explain the lack of material representing their families.
ECOLOGICAL NOTES ON THE METASEQUOIA AREA
Shui-sa-pa, as mentioned above, is at an elevation of 1050 meters, in a flat-floored valley or basin in a mountainous plateau or tableland region located about 60 km. southeast of the Yangtze River in the Li-chuan District, southwestern Hupeh Province (lat. 30'10' N., long. 108'35' E.). The Chi-yao Mountains (Permian limestone) attain an altitude of 1500 meters and bound the basin at an oblique angle on the northwest; mountains of another range, the Fu-pao-shan (Jurassic sandstone) reach about 1400 meters and bound the basin on the east.
The main stream of the basin occupies a bed of sandstone for most of its course and flows in a southerly direction for about 25 km. from its headwaters, then "roughly eastward in an S-curve for about 20 km. to the point where it disappears underground at the foot of limestone cliffs 200 m. high" (Chu & Cooper, 1950, p. 264) that form the southern boundary of the basin. Surrounded by mountains or high bluffs, the basin is effectively isolated.
The basin floor in the vicinity of Shui-sa-pa is between 300 and 600 meters wide, has mostly sandstone-derived soils, and is extensively used for the cultivation of rice, while the slopes of the surrounding mountains are "cut by ravines covered by semi-natural vegetation" (Chu & Cooper, 1950, p. 264).
The available climatic data for the metasequoia area have been discussed by Chu & Cooper (1950), who suggest that the region is similar with respect to temperature and total precipitation to the Coastal Plain of Georgia in the southeastern United States. However, in contrast to those of the latter area, the winters in the metasequoia area are relatively dry, while the summers are wet since most of the precipitation falls during the months of June, July, and August.
CHARACTERISTICS OF THE METASEQUOIA FLORA
As I worked with the specimens from the metasequoia area, the characteristics of the flora became increasingly clear. The most striking aspects that became evident relate to the diversity of forms seen in the large number of families and genera, the nature of the few prominent families and large genera in the area, the preponderance of woody species, the proportions of deciduous and evergreen species, the large number of endemic Chinese taxa represented, and the absence of weedy and high mountain species.
DIVERSITY. The diversity of the taxa is the most striking feature of the metasequoia flora. In an area of approximately 800 square kilometers, documentation exists for the presence of at least 550 species of vascular
[p. 48]
plants in 301 genera of 127 families, although the Gramineae and Palmae, as mentioned above, are inadequately represented in the collections studied. Thirty-one species are in 24 genera of 15 families of Pteridophyta; 18 species in 15 genera of six families in three classes of Gymnospermae; 357 species in 167 genera of 94 families of Dicotyledoneae; and 111 species in 66 genera of 12 families of Monocotyledoneae.
Three-fourths of the specimens are known to have been collected from the immediate vicinity of the metasequoia grove, which is a very small area situated at Shui-sa-pa. It should be mentioned that during Hwa's 1947 trip, he collected before reaching Shui-sa-pa. Some of the species that are represented by his numbers below 180 and that are not duplicated by his higher numbers are species that do not occur in the immediate vicinity of Shui-sa-pa. In the enumeration of species, there are at least 15 that do not grow in the metasequoia community, including Larix potaninii, Juniperus squamata, Eucommia ulmoides, Liriodendron chinense, Lindera viridis, Pittosporum glabratum, Euodia mellifolia, Zanthoxylum simulans, Rhus potaninii, Acer longipes, A. mono, Cercis chinensis, Ilex ficoidea, Elaeagnus henryi, and Alangium handelii. Some of these species may have been gathered from cultivated plants, since Eucommia has medicinal properties, Zanthoxylum is used as a spice, Cercis is an ornamental, and all are frequently cultivated.
The diversity of the flora is more pronounced when one considers the large number of families and genera that are represented by a small number of taxa. Seventy-three of the 127 families (almost 58%) in the area are represented by a single genus. In addition, there are 15 more families (almost 12%) represented by only two genera, and 12 others (ca. 10%) represented by three. Of the 301 genera, 202 (69%) are represented by a single species, 41 (14%) by two species, and 24 (ca. 8%) by three. The overwhelming number of families and genera represented by one or a few genera and species is indicative of the morphological distinctiveness of the components of the flora and probably reflects their history and that of the metasequoia area.
WELL-REPRESENTED FAMILIES AND GENERA. There are a few well-represented families and genera in the metasequoia area. The largest family is the Rosaceae, with 50 species in 16 genera. Additional large families include the Fagaceae (5/29) [2], Lauraceae (7/28), Ericaceae (5/19), and Theaceae, (4/10); the genera/species ratios of these families vary from 2.5 to 5.8.
Other families, including the Leguminosae (14/17), Liliaceae (8/10), Araliaceae (7/10), Compositae (8/9), Cyperaceae (7/8), Pinaceae (6/7), Euphorbiaceae (6/6), Polypodiaceae (5/7) and Labiatae (5/6), have relatively large numbers of genera, yet the numbers of species are comparatively small. These families have low genera/species ratios ranging between 1 and 1.4.
All the well-represented families with high genera/species ratios contain small genera. Examples include the Fagaceae (Castanea (2), [3] Fagus (2));
[2] The numbers in parentheses represent the number of genera and species, respectively, in the families concerned.
[3] The number in parentheses represents the number of species of the genus in the metasequoia flora.
[p. 49]
Lauraceae (Actinodaphne (1), Neolitsea (2), Sassafras (1)); Rosaceae (Agrimonia (1), Chaenomeles (1), Kerria (1), Stephanandra (1), Stravaesia (1)); Theaceae (Eurya (1), Ternstroemia (1)); and Ericaceae (Lyonia (1), Pieris (1)).
The genus with the greatest diversity in the metasequoia area is Quercus, which is represented by, 13 species. Other well-represented genera include Lindera (12), Prunus (12), Acer (11), Ilex (11), Viburnum (11), Rhododendron (9), Litsea (7), Salix (6), Rubus (6), Vaccinium (6), and Symplocos (6). Some of these genera are represented by species belonging to several subgeneric taxa; for example, in Ilex the eleven species belong to six different subgenera.
WOODY VS. HERBACEOUS SPECIES. The metasequoia flora has a preponderance of trees, shrubs, woody lianas, and perennial herbs. Woody species make up 76.4% of the 550 species, and 76.6% of the herbaceous species are perennials. It is possible, however, that herbs were under-collected (see above). It is noteworthy that the flora includes three parasites, one woody (Taxillus sutchuensis, Loranthaceae) and two herbaceous (Cuscuta chinensis, Convolvulaceae, and Protolirion sinii, Liliaceae).
DECIDUOUS VS. EVERGREEN SPECIES. Approximately 56 percent of the species associated with metasequoia are deciduous, while the remaining 44 percent are evergreen. The distribution of evergreen and deciduous species in the families is variable. In such families as the Juglandaceae and Betulaceae, all species are deciduous; in others a few species are deciduous, while the majority is evergreen. For example, in the Aquifoliaceae three species belonging to three different subgenera of Ilex are deciduous, yet eight others belonging to five subgenera are evergreen. The condition in the Pinaceae and Lauraceae is similar. In still other families (for example, the Taxaceae and Theaceae), all species are evergreen.
ENDEMIC GENERA AND SPECIES. The metasequoia flora is characterized by the presence of a high proportion of unusual plants found only in China. Some of the species, like Metasequoia glyptostroboides, belong to monotypic genera restricted to Central China; Eucommia ulmoides and Tetracentron sinense are additional examples. Other species, including Decaisnea fargesii, Dichroea febrifuga, Stranvaesia davidiana, Tapiscia sinensis, Idesia polycarpa, Poliothyrsis sinensis, Nothopanax davidii, and Serissa foetida, are endemic to China and belong to oligotypic genera. The degree of endemism and the ranges of the endemic genera are well illustrated by one-third of the gymnosperms that occur naturally with metasequoia (see below).
PAUCITY OF WEEDY AND HIQH ALTITUDE SPECIES. Having lived eight years in the Chengtu Plain and having traveled and botanized in the high mountains known as the rain-screen of the Sino-Tibetan border, I find the absence or paucity of various species a very striking character of the metasequoia flora. (1) Alnus crematogyne Burkill (Betulaceae) and Camptotheca acuminata Decaisne (Nyssaceae) are elsewhere common and conspicuous along streams and by the banks of rice fields. (2) There are few species of the families Chenopodia-
[p. 50]
ceae, Amaranthaceae, Aizoaceae, and Caryophyllaceae, which include many weedy species. (3) The numerous species of Abies Miller and Picea A. Dietrich, which form extensive forests in the mountainous regions of China, were not represented in the five collections under study from the metasequoia area. Perhaps the metasequoia area is not high and cool enough to support species of these two genera.
GYMNOSPERMS OF THE METASEQUOIA AREA
Seventeen species belonging to fourteen genera and six families of Gymnospermae occur naturally with Metasequoia glyptostroboides. Data on the distributional patterns and paleobotanical records of each genus might advance our understanding of the history, evolution, and phytogeographic significance of the metasequoia flora. With this end in view, information concerning these genera has been brought together below, and the data published by Florin (1963) have been heavily drawn upon.
The classes and orders to which the gymnospermous families belong, the genera in each family, the number of species in each genus in the metasequoia flora, and the present geographic range of the genera are given in TABLE 1. All of these genera are known from the fossil record.
ANTIQUITY. The genus Metasequoia Miki was first described from Lower Pliocene fossil material from Japan (Miki, 1941). When M. glyptostroboides was discovered as an extant plant, the botanists who described it and those who were associated with its introduction and distribution often referred to it as a "living fossil." The metasequoia area supports more than a dozen genera of gymnosperms (TABLE 1), and many of the species belong to monotypic or oligotypic genera now growing at low and middle latitudes in China. As evidenced by the fossil record, however, these genera once had much greater ranges in the Northern Hemisphere, and some of them have a longer geologic history than Metasequoia. For example, fossil remains of Pseudolarix, Cupressus, and Torreya have been recorded from Mesozoic deposits and are among the oldest known genera of conifers and taxads. As is well known, the ginkgos are among the oldest known gymnosperms, dating from the early Mesozoic (Andrews, 1961).
MORPHOLOGICAL DISTINCTIVENESS. Including Metasequoia glyptostroboides and Ginkgo biloba, the eighteen species of gymnosperms in the metasequoia area are distributed in fifteen genera, six families, and three classes. With the exception of Pinus and Juniperus, these genera are represented by single species in the metasequoia flora. In both Pinus and Juniperus the species belong to distinct subgeneric groups: the two species of Pinus to subg. PINUS (Diploxylon) and subg. STROBUS (Haploxylon), and the three species of Juniperus to sects. JUNIPERUS (Oxycedrus) and SABINA, the latter a taxon often recognized as a distinct genus.
Obviously, the gymnosperms of the metasequoia area represent many isolated evolutionary lines. Each species has a distinctive genotypic as well
[p. 51]
as phenotypic expression, and there is little chance for development of newer types through hybridization. Under favorable growth conditions they can survive, and they have persisted from the Mesozoic or the Cenozoic Era to the present day.
DISTRIBUTIONAL PATTERNS. The present geographic ranges of the gymnosperm genera that occur with metasequoia exhibit six distinctive distributional patterns. These are: (1) genera widespread in the Northern Hemisphere, including one genus (Juniperus) with a southward disjunction in the lake-mountain area of eastern equatorial Africa, and another (Taxus) with a range extending from southeastern continental Asia to the Philippines and the Celebes; (2) genera with eastern Asian and eastern North American disjunct distribution; (3) one genus (Pseudotsuga) with eastern Asian and western North American disjunct distribution; (4) one genus (Cupressus) with Sino-Himalayan-Mediterranean and western North and Central American distribution; (5) one genus (Cephalotaxus) with Sino-Japanese distribution; and (6) five genera, including Metasequoia, endemic to China at low and middle latitudes. These patterns clearly suggest that the gymnosperms of the metasequoia area have had different evolutionary histories and divergent adaptive radiations.
GENERA OF WIDESPREAD DISTRIBUTION. Four genera (Pinus and Larix, of the Pinaceae; Juniperus, of the Cupressaceae; and Taxus, of the Taxaceae) represented in the metasequoia flora are widespread in the Northern Hemisphere. The number of extant species in each of these genera is disputable; Pinus and Juniperus both contain many species that are distinguished by technical characters. However, the ranges of the sections or subgenera into which these genera have been divided are phytogeographically significant.
Pinus. The large genus Pinus has been subdivided into subg. PINUS and subg. STROBUS. Florin (1963, pp. 253-256, figs. 57, 58) presented an illustrated account of the distribution of the extant and fossil species. Both subgenera, as noted above, are represented in the metasequoia flora, with P. massoniana representing subg. PINUS and P. armandii typifying subg. STROBUS.
Larix. Like Metasequoia, Ginkgo, and Pseudolarix, the genus Larix is composed of deciduous species. It has a disjunct distribution in the Northern Hemisphere (cf. Florin, 1963, fig. 55) and has four centers of diversity: the northern Rocky Mountains of western North America; eastern Siberia-North China, Korea, and Japan; the interior of western and southwestern China and the adjacent eastern Himalayan area in northern Burma and northern India; and eastern Europe. In China, areas of diversity appear to be the Meridional Ranges on the Sino-Tibetan border, where four species are sympatric. Undoubtedly due to high altitude and sufficient precipitation, the species of this area extend the generic range ten to twenty degrees southward. However, Larix does not occur in southeastern China, as is stated by Florin (1963, p. 248).
[p. 53]
Juniperus. The genus Juniperus is large both in number of species and in the extent of its geographic range. Both sections of the genus accepted by Florin (1963, pp. 227-232, figs. 43, 44) are represented in the metasequoia flora, with J. formosana in sect. JUNIPERUS and J. chinensis and J. squamata in sect. SABINA.
Species of Juniperus are adapted to habitats in exposed areas with poor and often dry soils, and they often grow on barren rocky cliffs and windy ridges where seasonal and daily climatic fluctuations are extreme. The ranges of the extant species indicate that the genus contains both widespread and restricted species. Juniperus communis L. is widespread; its numerous varieties have a circumboreal distribution. Apparently, there are several centers where species are concentrated, i.e.. southwestern North America, the Mediterranean area, Central Asia, and the Sino-Himalayan-Japanese region.
Juniperus occurs in the Northern Hemisphere at middle and high latitudes, and at high altitudes in low latitudes. The generic range is extended southward by species of sect. SABINA in both Central America and eastern Africa. There is no evidence that the genus is native to subtropical southeastern China and adjacent Indochina. Juniperus formosana, however, occurs in the high mountains of Taiwan, as well as on the mainland in Hupeh, Szechwan, Yunnan, and Kweichow provinces.
Taxus. The genus Taxus exhibits a widespread but discontinuous distribution (cf. Florin, 1963, fig. 61). In contrast to Juniperus, Taxus has fewer species, and the specimens examined give no evidence of overlapping ranges except in the Sino-Himalayan region. When the specimens from this area are studied critically, the situation may be clarified.
One of the interesting features of the distribution of Taxus is that the species in Europe and North America grow at higher latitudes than do the Asian taxa. It is noteworthy that T. wallichiana Zucc. extends from the Meridional Ranges of southwestern China, westward to northern Burma, Nepal, and northern India, and southeastward to Annam in Indochina, Luzon in the Philippines, and then southward to the Celebes.
EASTERN ASIAN - EASTERN AND WESTERN NORTH AMERICAN DISJUNCTIONS. Two genera represented in the metasequoia flora exhibit eastern Asian - eastern and western North American disjunct distributions, and in both instances closely related species pairs occur on the two continents.
Tsuga. The genus Tsuga is of moderate size with about twelve species. Florin has carefully illustrated and explained the distribution of the fossil and extant species, but it should be noted that in each area of the disjunct generic range (cf. Florin, 1963, fig. 53) there are two or more species, all with overlapping ranges. The Sino-Himalayan region appears to be the present center of diversity for the genus.
The eastern Asian - eastern and western North American disjunction pattern in Tsuga is expressed by paired species in sects. TSUGA and HESPEROPEUCE. The closely related species in sect. TSUGA are T. chinensis, of Central and western China, and T. caroliniana Engelm., of Virginia, North and South
[p. 54]
Carolina, Tennessee, and Georgia; in sect. HESPEROPEUCE, T. longibracteata Cheng, from the mountainous areas of Kweichow and northern Kwangtung provinces, and T. mertensiana (Bong.) Sarg., widespread in western North America.
Torreya. A small genus of four to six species, Torreya occupies a limited geographic range (cf. Florin, 1963, fig. 63). It should be noted that the specimens I have examined have been identified as belonging to six species, three from mainland China, one from Japan, and two from the United States. The species look very similar and have been distinguished from one another by the color and pubescence of the twigs, the length of the leaves, the size and shape of the seeds, and the depth of the surface folding of the ruminate "endosperm."
In Torreya the disjunct distribution in eastern and western North America and in eastern Asia is expressed by pairs of species, a fact that was recognized by Rehder and Wilson as early as 1914. They commented (Pl. Wils. 2: 7. 1914) that "it is worthy of note that the eastern American T. taxifolia . . . agrees with T. grandis [of China] . . . and that the western American T. californica . . . agrees with T. nucifera [of Japan]." At that time T. jackii Chun of Chekiang in eastern China was not known - there is an even more prominent affinity between it and T. californica Torrey. It is worthy of note that with the exception of T. grandis and T. jackii, comparatively distantly related species that both occur in Chekiang, the ranges of Torreya species do not overlap. Each species is restricted to an area that is small, particularly so in North America.
EASTERN ASIAN - WESTERN NORTH AMERICAN DISJUNCTIONS. Extant species of Pseudotsuga are distributed disjunctly in eastern Asia (five species) and western North America (two species) (cf. Florin, 1963, fig. 52). Pseudotsuga menziesii (Mirbel) Franco is widespread in western North America, occurring in the Rocky Mountains and the Coastal Ranges, extending from Alberta and British Columbia in Canada southward through Montana, Idaho, and Washington, to Texas, Arizona, and California in the United States, and into Coahuila, Nuevo Le6n, and Sonora in Mexico. The second North American species, P. macrocarpa (Torrey) Mayr, is confined to southwestern California.
In eastern Asia Pseudotsuga occupies a broken and scattered range, with P. japonica (Shiras.) Beiss. confined to the islands of Honshu and Shikoku of Japan; P. wilsoniana Hayata restricted to the mountains of Taiwan; P. gausenii Flous endemic to Chekiang and southern Anhwei; P. sinensis confined to the metasequoia area and eastern Yunnan; and P. forestii Craib occurring only in the high mountainous area of northwestern Yunnan and adjacent Sikiang. The genus may once have been more widely distributed in China, the present distribution having been brought about through the destruction of the natural forest by the conversion of woodlands and open hills into terraced fields. All the described Chinese species appear to be very similar, and future study may indicate that they represent geographic variants of a single widespread species. It should also be mentioned that paleobotanical material of Pseudotsuga has been found within or close to the ranges of
[p. 55]
the extant species, a situation unlike that found in the other small gymnosperm genera occurring naturally with metasequoia.
The floristic relationships between eastern Asia and western North America, as exemplified by Pseudotsuga, have received less attention from phytogeographers than those between eastern Asia and eastern North America. Nevertheless, this pattern of disjunction is common among the angiosperms. Several genera, predominantly herbaceous perennials and shrubs, are good examples of taxa confined to these regions: Lysichiton Schott (Araceae); Lloydia Salisb. (Liliaceae); Lithocarpus Blume (Fagaceace); Eurotia Adanson (Chenopodiaceae); Mahonia Nuttall and Achlys A. P. DC. (Berberidaceae); Parrya R. Br. and Smelowskia C. A. Meyer (Cruciferae); Chamaerhodos Bunge (Rosaceae); Chamaesaracha A. Gray (Solanaceae); and Boschniakia C. A. Meyer (Orobanchaceae).
SINO-HIMALAYAN, MEDITERRANEAN, AND WESTERN NORTH AND CENTRAL AMERICAN DISJUNCTION. The present range of the genus Cupressus illustrates a Sino-Himalayan, Mediterranean, and western North and Central American disjunct distributional pattern (cf. Florin, 1963, fig. 38). Species of the genus are adapted to arid habitats, and the main area of differentiation appears to occur in California, followed in importance by the Sino-Tibetan border area. Eleven species, most of them localized endemics, occur in California, while there are four sympatric species in the Sino-Tibetan border area.
Species of Cupressus generally grow on rocky outcrops, coastal cliffs, or ridges, in canyons, and on sunny slopes of dry hills in chaparral; in each of its centers of distribution there is at least one widespread species. Thus, C. arizonica Greene is widespread in the southwestern United States and northern Mexico; C. lusitanica Miller grows widely in Mexico, and in Guatemala and El Salvador in Central America; and C. torulosa D. Don occupies a wide range in the Sino-Himalayan region. In the Mediterranean area C. sempervirens L. extends from Asia Minor and Iran westward to Morocco and occurs disjunctly in Algeria. Elsewhere in eastern and southeastern Asia Cupressus is absent from the floras of Japan, Taiwan, Indochina, and Kwangtung and Kwangsi.
SINO-HIMALAYAN - JAPANESE DISTRIBUTION. The present distribution of Cephalotaxus is confined to the Sino-Himalayan-Japanese region (MAP 2; see also Florin, 1963, fig. 60). Taxonomists differ in their interpretation of the number of species comprising the genus. Rehder (1940, 1949) recognized three (C. harringtonia, C. fortunei, and C. oliveri) and has been generally followed by recent authors including Ohwi in his Flora of Japan (1965). I have also followed Rehder in this study. All three species occur in China: two are widely distributed, with C. harringtonia (Forbes) K. Koch and its three varieties (vars. harringtonia, drupacea (Seib. [sic] & Zucc.) Koidzumi, and sinensis (Rehder & Wilson) Rehder) occupying the periphery of the generic range (MAP 9, H, D, S, respectively), and C. fortunei occurring in the metasequoia area
[4] Distribution maps have been plotted on the basis of specimens deposited in the herbaria of the Arnold Arboretum and Gray Herbarium.
[p. 56]
as well as in the warmer provinces of China; the third, C. oliveri Masters, has a broken and scattered range and is restricted to western China. The Sino-Himalayan-Japanese distributional pattern represented by Cephalotaxus is very common in angiosperms - Platycarya Sieb. & Zucc., Houttuynia Thunb., Corylopsis Sieb. & Zucc., Euscaphis Sieb. & Zucc., Stachyurus Sieb. & Zucc., Helwingia Willd., Abelia R. Br., and Ligularia Cass. are a few of the many genera from a wide range of families that share similar geographic ranges.
ENDEMIC DISTRIBUTION. Six genera (40%) - and one-third of the species - of the gymnosperms occurring in the metasequoia flora are endemic to China. These genera are Ginkgo (Ginkgoaceae), Keteleeria and Pseudolarix (Pinaceae), and Cunninghamia, Metasequoia, and Taiwania (Taxodiaceae). It is noteworthy that the metasequoia area (lat. 30°30' N.) marks the northern limit of the range of all of them. The seeds of Ginkgo biloba have been used by the Chinese people as both food and medicine for centuries, and it would appear that all known trees owe their existence to man's effort, either through protection or by planting. As a result, its distribution is highly artificial and is not considered here. The remaining five genera exhibit four distributional patterns: widespread as in Cunninghamia and Keteleeria, bicentric as in Taiwania, strongly restricted (in the Lower Yangtze region) as in Pseudolarix, and extremely restricted as in Metasequoia itself.
Cunninghamia. Although the ranges of Cunninghamia and Keteleeria overlap in many places (MAPS 3, 4), the nature of the distribution of the two genera is different. Less morphological variation is evident in Cunninghamia (MAP 3; see also Florin, 1963, fig. 34), and only two species have been recognized. Cunninghamia konishii Hayata, distinguished by its short leaves and small cones, is confined to the mountains of Taiwan. Cunninghamia lanceolata is widely distributed; however, since it is a fast grower, is easily propagated, and is often cultivated for its wood, its widespread distribution is partially the result of man's activities.
According to Florin (1963, p. 215), Cunninghamia has been recorded from Oligocene and Miocene deposits from Central Europe, and from an Oligocene deposit from Oregon.
Keteleeria. The occurrence o f Keteleeria in China (MAP 4; see also Florin, 1963, fig. 51) is spontaneous; it is more abundant away from human habitation, however, and is found only in mountainous areas. Rare in eastern China, it is frequent in the high mountains of the Sino-Tibetan border, especially at altitudes of 2300-3300 meters.
Keteleeria appears to be phenotypically plastic, depending upon the age of the plants and the environmental conditions under which they are growing. Due to this variability, the genus has received varying taxonomic interpretations. In 1926 E. H. Wilson recognized one species with a bicentric distribution in Taiwan and western China, while Flous (1936) recognized nine species. The characters Flous used to distinguish these species are phenotypically variable and include leaf length and apex shape. In 1978 Cheng and Fu
[p. 58]
recognized eleven species, nine of them from China (including one from Hainan). Again, the characters employed are phenotypically variable: length of leaves, hairs or papillae on young branchlets, shape of cone scales, and characters of the stomatic bands along the leaf midrib.
Wilson studied the plants in the field in the mountains of both Taiwan and western China. He observed (1926, p. 53) that the leaves of the same species vary from "2 to 5 cm. and may be rounded, emarginate, obtuse or acute at apex. On young trees and adventitious branches of old trees, the leaves are usually sharply acute or spinescent. . . . In length the cones vary from 5 to 20 cm. . . . The bracts vary in shape somewhat according to age and size. . . ." Orr (1933a), after analyzing the western Chinese collections of Forrest, Maire, and others, decided that all of the specimens belong to one species, Keteleeria davidiana.
The taxonomic status of species recognized in the genus awaits biosystematic investigation. Data for assessing the taxonomic value of morphological characters heretofore used for distinguishing species are needed. The problems involved in understanding the Keteleeria complex exemplify the difficulties inherent in the study of the metasequoia flora. To illustrate these problems, the published data of Flous (1936) and Cheng and Fu (1978) have been correlated with herbarium material deposited at the Arnold Arboretum and the Gray Herbarium and plotted on MAP 4. Interpretation of these data indicates that Keteleeria is a genus of warm-temperate Central China and subtropical South China and adjacent northern Indochina, and that variation exits in the morphology of the cone scales and seeds. In the west and southwest cones with large, ovate-rhomboid middle scales are produced, and the seeds have wings broadest below the middle. Eastern and southeastern populations produce cones with suborbicular scales, and seeds with wings broadest above the center. In the middle of the range a diversity of intermediate forms occurs; the Taiwanese element is more closely related to these forms than to those of Yunnan or Fukien. It would also appear that the Nan-lin Range, especially the area along the border of Kwangsi and Kweichow, constitutes a center of diversification; in this region the ranges of several of the recognized elements (calcarea, cyclolepis, and pubescens) overlap.
According to Florin (1963, p. 242), Keteleeria had a wider range in the past than it does now. It is represented in fossil remains from Europe in deposits ranging from the Oligocene to the Pliocene; it is also known from the Oligocene and Miocene from the Pacific Northwest and from Pliocene deposits in Japan.
Pseudolarix. The genus Pseudolarix is monotypic, and its range is restricted to eastern and Central China. Specimens of Pseudolarix from the metasequoia area extend its range 400 kilometers westward from its previously known occurrence (MAP 5; see also Florin, 1963, fig. 56). Pseudolarix amabilis was first discovered at Ning-po on the coast of Chekiang Province. In 1910 Wilson found it at an altitude of 1300 meters on Lu-shan, Kiangsi Province; based on this collection, Rehder and Wilson wrote that "this remarkable tree reaches the western limits of its distribution on the Lushan Range . . . where it
[p. 60]
is rare, and no large trees remain" (Pl. Wils. 2: 22. 1914). In 1918 Handel-Mazzetti collected some specimens in Hsin-hua, Hunan Province, and these collections extended the range of the species 400 kilometers into the interior of China. Later, the gap between the Chekiang and Kiangsi localities was filled in by Steward's 1924 collection from Chu-hua-shan in Anhwei and by R. C. Ching's specimens, collected in 1925, from Huang-shan in the same province. Cheng and Fu (1978) reported its occurrence in southern Kiangsu in the north and in northern Fukien in the south.
According to Florin (1963, p. 250), Pseudolarix had a wide range in the Northern Hemisphere in geologic time, existing in western North America in the Oligocene, and in Europe and Japan as late as the Pliocene.
Taiwania. Taiwania is treated by most taxonomists as a monotypic genus comprising the polymorphic species T. cryptomerioides. However, Cheng and Fu (1978) recognized the mainland material from Yunnan, Kweichow, and the metasequoia area as T. flousiana Gaussen. Taiwania cryptomerioides was described in 1906, and for ten years it was regarded as endemic to Taiwan. In 1915 Handel-Mazzetti collected the species in northwestern Yunnan, and based on this material Wilson (1926, p. 59) commented that
[p. 62]
"the occurrence of Taiwania . . . and of Pseudotsuga wilsoniana in western China and on the mountains of Formosa and at no other place in between these widely separated regions is a remarkable fact in plant distribution." Later, Orr (1933) reported a collection (J. H. Lace 52) made in 1912 from Upper Burma. In 1978 Cheng and Fu recorded it from southeastern Kweichow. The specimens from southeastern Kweichow and the metasequoia area have partially filled the distributional gap between Yunnan and Taiwan.
Specimens in the herbarium of the Arnold Arboretum indicate that the species is rather frequent in the sparsely populated mountainous areas of northeastern Yunnan and the adjacent border areas with Szechwan, Sikiang, and Upper Burma (MAP 6; see also Florin, 1963, fig. 35). In these areas, it occurs at altitudes of 2100-3700 meters in mixed evergreen forests.
EVOLUTIONARY AND PHYTOGEOGRAPHIC IMPLICATIONS
The characteristics of the metasequoia flora and the distributional patterns exhibited by the gymnosperms growing in association with Metasequoia glyptostroboides have significant implications concerning phytogeography and the evolutionary history of the vegetation. The antiquity of the metasequoia
[p. 63]
flora, its status as a relict flora, its origins and probable migration, and its floristic affinities are discussed below.
ANTIQUITY. The vegetation of the metasequoia area is a living sample of a comparatively well-preserved ancient flora. As mentioned above, all the genera of gymnosperms that occur naturally in the area have fossil records in the Tertiary epochs, and some extend back into the Mesozoic era. Paleobotanical discoveries indicate, moreover, that many of the lineages of the angiosperms represented in the metasequoia flora can also be traced back to the Tertiary. Wolfe's work (1969, 1978) on the Neogene floristic and vegetational history of the Pacific Northwest and Miki's findings (1954) concerning the Pliocene flora of Japan corroborate this contention. Wolfe (1969) listed 108 genera as present in the fossil remains of northwestern North America from the early and middle Miocene epoch. Sixty-two of these genera (57.5%) are also represented in the extant metasequoia flora. In addition, 21 other genera (18.5%) occur elsewhere in China (TABLE 2). Some of these genera, such as Abies and Picea, are important elements of boreal or montane taiga, while others, such as Glyptostrobus Endl., Gordonia Ellis, and Exbucklandia R. W. Br., are elements of subtropical lowland vegetation or montane forests.
In 1954 Miki reported on the occurrence of fossil remains of Taiwania, as well as of 26 other species, in the Pliocene clay and lignite beds of Japan. Eighty percent of the genera occurring in these beds are represented in the metasequoia flora, including Cunninghamia, Keteleeria, Metasequoia, Pseudolarix, Taiwania, Tsuga, Buxus, Fagus, Hydrangea, Ilex, Juglans, Liquidambar, Quercus, Pterocarya, Schisandra, Spondias (= Choerospondias), Styrax, and Symplocos.
Based on these fossil occurrences, the question that immediately comes to mind is why so many genera and species have disappeared from western North America and Japan but have survived in China (and in the metasequoia area in particular). The answers lie largely in the relative amount of change in topography and climate that have occurred in these regions subsequent to the establishment of modern floras in the early Tertiary or late Mesozoic. The floras of the metasequoia area and the region to the south in China have been spared from the direct effects of the repeated Pleistocene continental glaciation, from the aridity created by the mountain building and continental uplift during the Miocene and Pliocene (which gradually turned forested regions into steppes and deserts as in areas of the same latitude in Tibet), and from the effects of local mountain glaciation and vulcanism (as in Japan).
Although shifts of the climatic belts during the glacial and interglacial ages of the Pleistocene were worldwide and contemporaneous, local conditions in Central China reduced the intensity of the effects of lower temperatures and perhaps also the increased precipitation; hence, no glaciers were formed. As shown in MAP 7, eastern Asia was not covered by continental ice sheets. High mountains to the west and northwest of the metasequoia area and, according to Lee (1939, pp. 376-399), a few lower mountains in the Lower Yangtze Valley to the east (eg. [sic], Lu Shan in Kiangsi, Huang Shan in Anhwei, and Tien-mu Shan in Chekiang) were marked by local glaciers. Geographically,
[p. 64]
the metasequoia area is situated in the Middle Yangtze region at middle latitude. During the glacial periods, it received a less pronounced increase in precipitation than did the mountainous areas mentioned above. Geologically, the continuous Tsinling Range of high mountains on the north, northwest, and west formed a natural "Great Wall" that shielded the area from the intense cold of the southward-shifting northern climatic zones. Moreover, the southwest-northeast orientation of the mountains of the metasequoia area permitted meridional circulation of warmer air. As a result of the combination of these factors, the vegetation of the metasequoia area was undoubtedly far less disrupted than the vegetation and floras of areas covered by continental glaciation in Europe and North America, and also less disrupted than the nearby vegetation of the Lower Yangtze Valley. While direct evidence on this point is unavailable, it may be that Metasequoia glyptostroboides and its associates survived the Pleistocene in situ in the metasequoia area.
[p. 65]
ORIGIN AND MIGRATION OF THE METASEQUOIA FLORA. After visiting the metasequoia area, Chaney (1948) concluded that the general aspect of the Metasequoia-hardwood assemblage is similar to that of the Taxodium-hardwood association of moist flood plains of the southeastern United States. For the early and middle Tertiary he reconstructed a profile of mild climate with summer rainfall and moderate temperature not regularly falling below freezing. Metasequoia and Taxodium were both widely distributed at high latitudes during the early Tertiary and occurred in western North America and northern Asia during the middle Tertiary. Chaney also advanced a theory with regard to the origin and migration of the Arcto-Tertiary metasequoia flora. "We conclude that this forest type had its origin in the north, in Eocene and Cretaceous time . . . and its southward migration during the Tertiary period." Chaney proposed this theory on the strength of the "deciduous habit of the dominant trees, including Metasequoia and Taxodium." He suggested that the length of the arctic night was the primary factor causing the deciduous habit.
Floristic analysis of the metasequoia flora and recent paleobotanical evidence do not support Chaney's theory. First, the deciduous habit is not predominant in the metasequoia flora since approximately 44 percent of the species are evergreen. Of the 18 species of gymnosperms in the area, only one-sixth are deciduous. These include Larix potaninii, Metasequoia glyptostroboides, and Pseudolarix amabilis, all of which grow in wet habitats and are hydrophilous. It would appear that the alternation of a dry winter season with a wet summer results in a fluctuation in the water supply that affects physiological activities, inducing the deciduous habit of these conifers. In the angiosperms, the texture of the leaves and the effects of dry winter on the plants are
[p. 66]
obviously the controlling factors governing the annual shedding of leaves in areas where frost never occurs. In the tropical monsoon forest of Hong Kong, I have observed two definite periods when leaves are shed. Species with thin, chartaceous leaves shed them after the beginning of the dry season and appear deciduous in winter. Species with coriaceous leaves drop them after the dry season as, or soon after, flower and leaf buds unfold in early spring. This latter group of species appears evergreen in winter. A similar phenomenon was observed in Chengtu in the Red Basin of Szechwan, where frost seldom occurs.
Second, Chaney's theory of the origin and direction of migration of the metasequoia flora is oversimplified. His assumption of a holarctic origin and southward migration of the flora was formulated on the basis of an insufficient number of species from the metasequoia area. He listed (1948, p. 510) only eight species of deciduous dicotyledonous angiosperms, four species of gymnosperms, and one palm. At the generic level, his list includes less than twenty-five percent of the genera of gymnosperms, and less than four percent of the angiosperms now documented as growing in the metasequoia area. Yet, based on this scant evidence, Chaney concluded that "this assemblage is essentially the same as that recorded in the fossil record of the Eocene from high latitudes. . . ."
Numerous recent paleobotanical discoveries have negated the concept of an Arcto-Tertiary geoflora with temperate broad-leaved deciduous forests occurring at high latitudes. To the contrary, the high percentages of large leaves with entire margins, palmate venation, and elongated apices occurring in the fossil assemblages in Europe, Japan, and the western United States from middle Eocene and Miocene deposits suggests the existence of tropical rain forest vegetation (Wolfe, 1978).
Although the fossil record of Miocene western North America is very incomplete, it is perhaps noteworthy that 39 genera in 20 families (TABLE 3) are found in the metasequoia area but not in the Miocene of western North America. All 39 genera of the 20 families included in TABLE 3 occur in the southeastern portion of the Asian mainland and in Malaysia and are elements of tropical or subtropical forests. Moreover, all attain the northern limits of their ranges in or beyond the metasequoia area, indicating a southern affinity for the metasequoia flora. Although the genera all contain a large number of species outside the metasequoia area, representatives of these genera grow and multiply naturally in the same area with Metasequoia glyptostroboides.
These southern elements have very complicated floristic affinities. Although a few, such as Michelia, Eriobotrya, Camellia, and Schima, are confined to the Asiatic mainland with only one or two species in Japan or the Philippines, most have ranges that extend far into the Pacific land areas. Reference to TABLE 3 will show that several genera are world wide in distribution, while others exhibit tropical Asian-Pacific and tropical American affinities; eighteen genera show African relationships. The eastern North American - southeastern Asian distribution pattern is illustrated by Lindera. the western North American - eastern Asian distribution by Photinia (Heteromeles M. Roemer).
[p. 67]
This analysis of the metasequoia flora serves to point out its complexity; it gives neither a clue to its place of origin nor an indication of the direction of past migration routes. The gymnosperms have several distributional patterns: those exhibited by Larix and Tsuga indicate northern affinities, while those of Cephalotaxus, Keteleeria, and Taiwania attest to southern affinites [sic]. The information derived from the various phytogeographic elements points to the dynamic nature of plant evolution and distribution - the various taxa in a small area remind one of the phenomenon of traffic congested in a rotary, where slow-moving vehicles of all makes and ages are gathered together. The flora of any given region is never static; the metasequoia flora provides a sample of an ancient flora growing today in an area where it may have survived in situ and offers an example of the ecological conditions under which such a flora may have been able to survive and evolve. That the metasequoia area is ecologically diverse is evidenced by the complexity of its flora.
AN ENUMERATION OF THE SPECIES
PTERIDOPHYTA
(Identified with the assistance of Professor R. M. Tryon)
LYCOPSIDA
Lycopodiaceae
Lycopodium complanatum L. Cheng & Hwa 1081.
Selaginellaceae
Selaginella uncinata (Desv.) Sprengel, Cheng & Hwa 987.
FILICES
Plagiogyriaceae
Plagiogyria stenoptera (Hance) Diels, Cheng & Hwa 1138.
Schizaeaceae
Lygodium japonicum (Thunb.) Sw. Cheng & Hwa 998.
Lindsaeaceae
Sphenomeris chusana (L.) Copel. (Stenoloma chusana (L.) Ching), Cheng & Hwa 794.
Pteridaceae
Pteris henryi Christ, Cheng & Hwa 916.
Pteris vittata L. Cheng & Hwa 877.
Sinopteridaceae
Onychium japonicum (Thunb.) Kunze, Cheng & Hwa 647.
Gymnogrammaceae
Coniogramma japonica (Thunb.) Diels, Cheng & Hwa 646.
[p. 70]
Adiantaceae
Adiantum pedatum L. Cheng & Hwa 846.
Athyriaceae
Diplazium chinense (Baker) C. Chr. Cheng & Hwa 583.
Diplazium japonicum (Thunb.) Bedd. Cheng & Hwa 580, 585.
Thelypteridaceae
Cyclosorus aridus (D. Don) Ching (Aspidium aridum D. Don), Cheng & Hwa 874.
Cyclosorus parasiticus (L.) Farwell, Cheng & Hwa 581.
Thelypteris decursivo-pinnata (van Hall) Ching, Cheng & Hwa 584, 586.
Onocleaceae
Matteuccia orientalis (Hooker) Trev. Cheng & Hwa 645, 876, 881.
Aspidiaceae
Cyrtomium fortunei J. Sm. Cheng & Hwa 693, 915.
Dryopteris fuscipes C. Chr. Cheng & Hwa 694, 695.
Polystichum acutidens Christ, Cheng & Hwa 960.
Polystichum tripteron (Kunze) Presl, Cheng & Hwa 696.
Polystichum tsus-simense (Hooker) J. Sm. Cheng & Hwa 699, 959.
Polypodiaceae
Arthromeris lehmannii (Mett.) Ching (Polypodium lehmannii Mett.), Cheng & Hwa 906.
Lepisorus pseudonudus Ching, Cheng & Hwa 924.
Microsorium fortunei (Moore) Ching (Drymaria fortunei Moore), Cheng & Hwa 698.
Microsorium hymenodes (Kunze) Ching (Polypodium hymenodes Kunze), Cheng & Hwa 932.
Neolepisorus phyllomanes (Christ) Ching (Polypodium phyllomanes Christ), Cheng & Hwa 933.
Polypodium nipponicum Mett. Cheng & Hwa 912.
Pyrrosia lingua (Thunb.) Farwell, Cheng & Hwa 697.
Pyrrosia sheareri (Baker) Ching, Cheng & Hwa 750.
SPERMATOPHYTA
GYMNOSPERMAE
Ginkgoaceae
Ginkgo biloba L. Gressitt 2412.
Pinaceae
Keteleeria davidiana (Bertrand) Beiss. Cheng & Hwa 1175, Hwa 1, 450.
Larix potaninii Batalin, Hwa 80.
Pinus armandii Franchet, Cheng & Hwa 1147, 1166, Gressitt 2536, Hwa 14.
Pinus massoniana Lambert, Cheng & Hwa 655, 825, 1098, 1165, 1178; Gressitt 2405 (sterile); Hwa 439.
Pseudolarix amabilis (Nelson) Rehder. Cheng & Hwa 1169; Hwa 466.
[p. 71]
Pseudotsuga sinensis Dode, Cheng & Hwa 1170, Hwa 22, 449.
Tsuga chinensis (Franchet) Pritzel, Hwa 176, 543.
Taxodiaceae
Cunninghamia lanceolata (Lamb.) Hooker, Cheng & Hwa 1164; Gressitt 2403 (sterile).
Metasequoia glyptostroboides Hu & Cheng, Cheng & Hwa 558, 571, 617, 808, 816, 851, 1141, 1163, 1168, Gressitt 2401, 2564; Hwa 209;219, 220, 296, 342, 351.
Taiwania cryptomerioides Hayata, Cheng & Hwa 1099, 1172; Hwa 443, 460, 480.
Cupressaceae
Cupressus funebris Endl. Cheng & Hwa 828; Djou 104; Gressitt 2404, 2504, 2506.
Juniperus chinensis L. Gressitt 2505; Hwa 253.
Juniperus formosana Hayata, Cheng & Hwa 1176, Gressitt 2535; Hwa 39, 150, 547.
Juniperus squamata Buch.-Ham. ex Lamb. Hwa 252.
Cephalotaxaceae
Cephalotaxus fortunei Hooker, Cheng & Hwa 725, 814A; Gressitt 2402 (sterile); Hwa 21 (sterile), 214A, 218, 284, 299, 302, 415, 422, 509.
Taxaceae
Taxus chinensis (Pilger) Rehder, Cheng & Hwa 1142, Hwa 27, 164, 229, 294.
Torreya grandis Fortune, Gressitt 2407 (sterile); Hwa 201, 210, 211, 241.
ANGIOSPERMAE
Dicotyledoneae
Saururaceae
Houttuynia cordata Thunb. Cheng & Hwa 641.
Chloranthaceae
Chloranthus henryi Hemsley, Cheng & Hwa s.n.
Salicaceae
Populus adenopoda Maxim. Hwa 70 (sterile).
Populus tremula L. var. davidiana Schneider, Cheng & Hwa 663.
Salix biondiana Seem. Hwa 394, 395.
Salix caprea L. Hwa 113, 242.
Salix ernestii Schneider, Gressitt 2465, 2516, 2546; Hwa 438.
Salix heterochroma Seem. Hwa 379 (sterile).
Salix longiflora Andersson, Cheng & Hwa 805.
Salix rosthornii Seem. Hwa 391, 392.
Juglandaceae
Juglans cathayensis Dode, Gressitt 2458, Hwa 505, 536.
Juglans regia L. Cheng & Hwa 834, Gressitt 2401, 2528, 2547; Hwa 403.
[p. 72]
Platycarya strobilacea Sieb. & Zucc. Cheng & Hwa 1117; Gressitt 2509; Hwa 161.
Pterocarya hupehensis Skan, Cheng & Hwa 594; Gressitt 2425.
Pterocarya paliurus Batalin, Cheng & Hwa 863, 880, Hwa 492.
Pterocarya stenoptera C. DC. Cheng & Hwa 865, 961; Gressitt 2530, Hwa 463.
Betulaceae
Betula albo-sinensis Burkill, Cheng & Hwa 1080.
Betula alnoides Buch.-Ham. ex D. Don, Gressitt 2412, 2522, 2566, Hwa 400.
Betula fargesii Franchet, Hwa 299.
Betula luminifera Winkler, Hwa 257.
Betula wilsoniana Schneider, Cheng & Hwa 1078; Hwa 25, 386.
Carpinus fargesii Franchet, Cheng & Hwa 728, 1051, 1102; Gressitt 2468; Hwa 63.
Carpinus sp., Cheng & Hwa 866, 950.
Corylus fargesii (Franchet) Schneider, Cheng & Hwa 745, Hwa 20, 212.
Ostrya multinervis Rehder, Cheng & Hwa 661, 1137, Hwa 128, 267, 324.
Fagaceae
Castanea henryi Rehder & Wilson, Cheng & Hwa 548, Hwa 6, 15.
Castanea mollissima Bl. Cheng & Hwa 556, 726, Hwa 5, 323.
Castanopsis chinensis Hance, Cheng & Hwa 1012.
Castanopsis eyrei (Champ. ex Bentham) Tutcher, Cheng & Hwa 1009, Hwa 42, 442, 456.
Castanopsis hystrix A. DC. Cheng & Hwa 1007.
Castanopsis sclerophylla (Lindley) Schottky, Hwa 420.
Castanopsis tibetana Hance, Cheng & Hwa 1043; Hwa 301, 486.
Fagus longipetiolata Seem. Gressitt 667, 2481; Hwa 36, 321.
Fagus lucida Rehder & Wilson, Cheng & Hwa 1058, 1174.
Lithocarpus glabra (Thunb.) Nakai, Cheng & Hwa 776, 1021.
Lithocarpus hancei Rehder, Cheng & Hwa 748, 1073; Hwa 116, 334, 440.
Lithocarpus henryi (Seem.) Rehder & Wilson, Cheng & Hwa 683, 1074, 1095; Hwa 336.
Quercus acutissima Carr. Cheng & Hwa 771; Hwa 79.
Quercus aliena Bl. Cheng & Hwa 1126; Hwa 76, 517.
Quercus cocciferoides Hand.-Mazz. Cheng & Hwa 1054, Hwa 175, 444.
Quercus delavayi Franchet, Cheng & Hwa 856, 944.
Quercus engleriana Seem. Cheng & Hwa 907, 951, 986; Hwa 169.
Quercus fabri Hance, Cheng & Hwa 739, Hwa 8.
Quercus glauca Thunb. Cheng & Hwa 733, 1000, 1128, Hwa 85, 479, 522.
Quercus glauca Thunb. var. gracilis Rehder & Wilson, Cheng & Hwa 774, Hwa 145.
Quercus nubium Hand.-Mazz. Cheng & Hwa 612, Hwa 333.
Quercus oxyodon Miq. Cheng & Hwa 854, 893; Gressitt 2447, Hwa 308, 361.
Quercus singuliflora (Lévl.) A. Camus, Cheng & Hwa 600, Hwa 167, 330.
Quercus spathulata Seem. Cheng & Hwa 1119, Hwa 86, 407, 519.
Quercus stewardiana A. Camus, Hwa 451.
Quercus variabilis Bl. Hwa 62.
Quercus sp., Cheng & Hwa 120, 681, 1079; Hwa 287, 493 (flowering material too young to determine).
[p. 73]
Ulmaceae
Celtis labilis Schneider, Cheng & Hwa 962, 1131; Gressitt 2529; Hwa 108, 289, 431.
Trema laevigata Hand.-Mazz. Hwa 429.
Ulmus bergmanniana Schneider, Hwa 419, 513.
Ulmus multinervis Cheng, Cheng & Hwa 1130 (syntype), 1153, Hwa 286, 406 (syntype), 435 (all specimens det. W. C. Cheng).
Ulmus prunifolia Cheng, Hwa 258, 549 (syntypes).
Zelkova schneideriana Hand.-Mazz. Cheng & Hwa 798, 1136.
Eucommiaceae
Eucommia ulmoides Oliver Hwa 77.
Moraceae
Broussonetia papyrifera (L.) Vent. Gressitt 2543; Hwa 413, 474, 475.
Cudrania tricuspidata (Carr.) Bur. Cheng & Hwa 830, 943; Gressitt 2407, 2453, 2498, 2511; Hwa 467.
Ficus heteromorpha Hemsley, Gressitt 2561; Hwa 54, 138.
Ficus lacor Buch.-Ham. ex D. Don, Cheng & Hwa 1180.
Ficus sarmentosa Buch.-Ham. ex D. Don, Cheng & Hwa 709.
Ficus stapfii Lévl. Cheng & Hwa 1120; Hwa 411.
Morus cathayana Hemsley, Gressitt 2557; Hwa 326, 508, 514.
Urticaceae
Chamabainia cuspidata Wight, Cheng & Hwa 598, 843.
Elatostema sessile Forster, Cheng & Hwa 839.
Pilea notata Wright, Cheng & Hwa 872.
Loranthaceae
Taxillus sutchuenensis (Lecomte) Danser, Cheng & Hwa 939 (det. B. A. Razi).
Polygonaceae
Antenoron neofiliforme (Nakai) Hara, Cheng & Hwa 692.
Polygonum cuspidatum Sieb. & Zucc. Cheng & Hwa 716.
Polygonum multiflorum Thunb. Cheng & Hwa 966.
Polygonum perfoliatum L. Cheng & Hwa 976.
Polygonum runcinatum Buch.-Ham. ex D. Don, Cheng & Hwa 958.
Tetracentraceae
Tetracentron sinense Oliver, Cheng & Hwa 1067.
Cercidiphyllaceae
Cercidiphyllum japonicum Sieb. & Zucc. var. sinense Rehder & Wilson, Cheng & Hwa 562.
Ranunculaceae
Anemone hupehensis Lemoine, Gressitt 2551.
Clematis meyeniana Walp. Hwa 295.
Clematis montana Buch.-Ham. ex A. P. DC. Cheng & Hwa 627.
Clematis spooneri Rehder & Wilson var. subglabra S. Y. Hu, var. nov.
[p. 74]
A typo differt in foliolis praeter nervas glabris, foliolis lateralibus subsessilibus; pedunculis pilosis; achenis glabris.
Cheng & Hwa 1075 (holotype, A).
Clematis spooneri Rehder & Wilson is characterized by trifoliolate leaves with firm leaflets, the terminal ones elliptic, 5-7 cm. long, 2.5-4 cm. wide, with 2 or 3 large teeth on each side, the lateral leaflets ovate with oblique bases, 3.5-6.5 cm. long, 1.5-3.5 cm. wide, with 1 or 2 teeth on each side; and by elongated fruiting peduncles 1 1/2 - 2 1/2 times longer than the petioles. The species is closely related to C. chrysocoma Franchet, the, leaves of which are densely covered by coarse golden hairs. The leaves of C. spooneri var. spooneri are densely pilose, while those of C. spooneri var. subglabra are sparsely pilose on the primary nerves but otherwise glabrous.
Coptis chinensis Franchet, Cheng & Hwa 657.
Trollius yunnanensis Franchet, Cheng & Hwa 871.
Lardizabalaceae
Decaisnea fargesii Franchet, Cheng & Hwa 654; Gressitt 2438,
Berberidaceae
Berberis fallax Schneider, Cheng & Hwa 1158.
Berberis julianae Schneider, Hwa 157.
Menispermaceae
Cocculus orbiculatus (L.) A. P. DC. Cheng & Hwa 995.
Magnoliaceae
Liriodendron chinense (Hemsley) Sargent, Hwa 162.
Magnolia denudata Desrouss. Cheng & Hwa 1049, 1066, Hwa 65, 104, 213.
Magnolia officinalis Rehder & Wilson, Cheng & Hwa 1160, Hwa 38, 290, 404.
Magnolia sprengeri Pampan. Cheng & Hwa 1171; Hwa 81, 276, 528,
Michelia floribunda Finet & Gagnep. var. hupehensis Cheng, Cheng & Hwa 688, 1005 (isotype; det. W. C. Cheng); Hwa 317, 503.
Michelia martinii (Lévl.) Dandy, Hwa 411.
Michelia wilsonii Finet & Gagnep. Cheng & Hwa 889, 938, Hwa 357, 502.
Schisandraceae
Kadsura longipedunculata Finet & Gagnep. Cheng & Hwa 650.
Schisandra sphenanthera Rehder & Wilson, Cheng & Hwa 823, 825, Gressitt 2444.
Illiciaceae
Illicium brevistylum A. C. Sm. Cheng & Hwa 614.
Illicium lanceolatum A. C. Sm. Cheng & Hwa 936, Hwa 23, 154, 304, 551.
Calycanthaceae
Chimonanthus praecox (L.) Link, Cheng & Hwa 921.
Lauraceae
Actinodaphne cupularis (Hemsley) Gamble, Hwa 436, 483.
[p. 75]
Cinnamomum inunctum Nees var. albosericeum Gamble, Cheng & Hwa 782, Hwa 117.
Cinnamomum mairei Lévl. Cheng & Hwa 1028; Hwa 410, 497.
Cinnamomum platyphyllum (Diels) Allen, Cheng & Hwa 1010, Hwa 484, 506.
Lindera cercidifolia Hemsley, Cheng & Hwa 1108.
Lindera communis Hemsley, Hwa 34, 165, 386 (carpellate plant in flower; matches isotype in all important characters except that leaves are glabrous beneath), 515.
Lindera communis Hemsley var. esquirolii (Lévl.) S. Y. Hu, comb. nov.
Litsea esquirolii Lévl. in Repert. Sp. Nov. 9: 459. 1911.
Benzoin commune (Hemsley) Rehder in Jour. Arnold Arb. 1: 144. 1919, pro parte.
Rehder in 1919 considered Lindera communis Hemsley and Litsea esquirolii Lévl. to be synonymous. The holotype of L. esquirolii was collected outside Kwei-yang in Kweichow Province (Esquirol 372). It represents a taxon with leaves having a rufous lower surface, a densely gold-pubescent midrib, and primary lateral nerves. I have examined eight sheets of the syntypes of Lindera communis (Henry 1204, 1207, 1523, 2662, 3413, 3413B, 4551), all from western Hupeh. All these specimens have a glaucous lower leaf surface and are sparingly and very finely pubescent on the midrib and lateral nerves. The taxon with a rufous lower leaf surface and densely pubescent veins and twigs has a more southerly distribution than the typical variety. Additional specimens of var. esquirolii examined are Cheng & Hwa 743, 858, 1050, and Hwa 418, 516.
Lindera fruticosa Hemsley, Cheng & Hwa 576, 636, Gressitt 2572.
Lindera glauca Sieb. & Zucc. Cheng & Hwa 575, Gressitt 2487, 2549; Hwa 19, 100.
Lindera hemsleyana (Diels) Allen, Hwa 307, 408, 414, 507.
Lindera ichangensis Gamble, Djou 110; Gressitt 2571.
Lindera megaphylla Hemsley, Cheng & Hwa 857, Hwa 103.
Lindera stewardiana Allen, Cheng & Hwa 779.
Lindera subcaudata (Merr.) Merr. Cheng & Hwa 619, 963; Hwa 35, 235.
Lindera umbellata Thunb. Cheng & Hwa 952 (too immature to be identified with certainty).
Lindera sp., Hwa 88.
Litsea chunii Cheng, Hwa 246.
Litsea cubeba (Lour.) Pers. Cheng & Hwa 797, 885, 1011; Djou 125, Hwa 215, 233.
Litsea elongata (Nees) Hooker f. Cheng & Hwa 672, 895, 1047, 1106; Hwa 109, 266.
Litsea glutinosa (Lour.) C. B. Robinson, Cheng & Hwa 815.
Litsea pungens Hemsley, Hwa 259, 264, 313.
Litsea verticillata Hance, Hwa 371 (sterile; identification not certain).
Machilus chinensis (Champ. ex Bentham) Hemsley, Hwa 141, 468.
Machilus ichangensis Rehder & Wilson, Cheng & Hwa 886.
Machilus lichuanensis Cheng ex Li, Cheng & Hwa 1017 (syntype); Hwa 476 (syntype; det. W. C. Cheng).
Neolitsea sieboldii (O. Kuntze) Nakai, Cheng & Hwa 668.
Neolitsea viridis Cheng & S. Y. Hu, sp. nov.
Arbor sempervirens, ramulis glabris; foliis subcoriaceis, ellipticis, 4.5-7 cm. longis, 1.2-2.5 cm. latis, acuminatis, ad basin acutis, supra prominente
[p. 76]
reticulatis, glabris, viridis, subtus foveolatis, sub lente pilosis, penninervis, nervis 9- vel 10-jugatis, subalternatis; petiolis 10-14 mm. longis, glabris rugosisque; inflorescentiis fasciculatis, axillaribus, subsessilibus; floribus masculinis pedicellis dense ferruginoso-hirsutis, 2 mm. longis; perianthis 4-lobatis, tubis utrinque ferruginoso-hirsutis, lobis ovatis, ciliatis, 3 mm. longis, 1.5 mm. latis, extus pubescentibus, intus glabris; staminibus 6, exsertis, filamentis 4 mm. longis, glabris, 2 raro 1 interioribus bi-glandulosis, glandulis breviter stipitatis, antheris magnis; ovariis rudimentalibus, 1-2 mm. longis, glabris; floribus femineis et fructibus ignotis.
HUPEH: Li-chuan District, Tuan-pao-hsiang, C. T. Hwa 125 (holotype, A).
This collection was distributed as a new species of Litsea. After having checked all the collections of the genera Litsea and Neolitsea in the herbarium of the Arnold Arboretum and having dissected the flowers, I decided to place it in Neolitsea. The species is closely related to N. wushanica (Chun) Merr., which differs in having glaucous lower leaf-surfaces, and "pedicellis sericeo-hirsutis." From N. pinninervis Yang & Huang, it differs in having glabrous filaments and rudimentary ovaries.
Sassafras tzumu (Hemsley) Hemsley, Cheng & Hwa 764; Djou 105, Gressitt 2440; Hwa 208.
Papaveraceae
Macleaya cordata (Willd.) R. Br. Cheng & Hwa 1021.
Saxifragaceae
Astilbe chinensis Franchet & Sav. Gressitt 2478.
Dichroa febrifuga Lour. var. glabra S. Y. Hu, var. nov.
A typo differt in ramis glabris; foliis minoribus, 5-11 cm. longis, 2-4 cm. latis, utrinque glabris; inflorescentiis, hypanthiis, calycibus, petalis et antheris omnino glabris.
Cheng & Hwa 587 (holotype, A).
Other specimens examined. HUPEH: En-shih, H. C. Chow 1881; Pa-tung, H. C. Chow 697 (a better specimen than the holotype; in flower), 1025. SZECHWAN: Chung Hsien, W. P. Fang 492; Nan-chuan, W. P. Fang 1246, 5720.
Dichroa febrifuga Lour. was described on the basis of Loureiro's collection from the vicinity of Canton, now deposited in the British Museum. Many specimens from this area, as well as from many other provinces in the warmer region of China, were examined. A wide range of variation exists in the size of the plants, the shape, texture, and dentation of the leaves, and the number of flowers per cluster. It appears that the nature of trichomes on the leaves and inflorescences is constant. A distinctive species with strigose-villose indumentum, D. longipila Merr., was described from the mountains on the border of Kwangtung and Kwangsi. Here a glabrous variety of D. febrifuga is reported from the mountains on the border of Hupeh and Szechwan provinces.
Hydrangea chinensis Maxim. Cheng & Hwa 884.
Hydrangea paniculata Sieb. Cheng & Hwa 1065.
Hydrangea strigosa Rehder, Cheng & Hwa 812; Gressitt 2437, 2464.
Hydrangea umbellata Rehder, Cheng & Hwa 1082.
Parnassia wrightiana Wall. var. flavida Franchet, Cheng & Hwa 778.
Philadelphus sericanthus Koehne, Cheng & Hwa 602,
[p. 77]
Pittosporaceae
Pittosporum adaphniphylloides Hu & Cheng, Hwa 268.
Pittosporum glabratum Lindley var. glabratum, Hwa 133.
Pittosporum glabratum Lindley var. neriifolium Rehder & Wilson, Hwa 340.
Pittosporum truncatum Pritzel, Hwa 405.
Hamamelidaceae
Corylopsis henryi Hemsley, Hwa 245.
Corylopsis platypetala Rehder & Wilson, Hwa 281, 283.
Corylopsis sinensis Hemsley, Cheng & Hwa 1057, Hwa 49.
Liquidambar formosana Hance, Cheng & Hwa 708; Gressitt 2415.
Loropetalum chinense Oliver, Cheng & Hwa 1024.
Sycopsis sinensis Oliver, Cheng & Hwa 747, 1179, Hwa 260, 271, 300, 542.
Rosaceae
Agrimonia pilosa Ledeb. Cheng & Hwa 658.
Chaenomeles cathayensis Hemsley, Hwa 292, 469.
Cotoneaster gracilis Rehder & Wilson, Hwa 387.
Cotoneaster hebephylla Diels, Cheng & Hwa 653, 1048.
Cotoneaster horizontalis Decaisne, Cheng & Hwa 781.
Cotoneaster salicifolia Franchet, Cheng & Hwa 940, Hwa 123.
Eriobotrya cavaleriei (Lévl.) Rehder, Hwa 146, 448, 588.
Eriobotrya deflexa Hemsley, Hwa 344.
Eriobotrya japonica (Thunb.) Lindley, Cheng & Hwa 1159; Hwa 526.
Kerria japonica DC. Cheng & Hwa 595; Hwa 251.
Malus halliana Koehne, Cheng & Hwa 1025; Hwa 279.
Malus prunifolia (Willd.) Borkhausen, Hwa 401.
Photinia amphidox (Schneider) Rehder & Wilson, Cheng & Hwa 1151; Hwa 122, 269.
Photinia beauverdiana Schneider, Cheng & Hwa 999, Hwa 94, 110, 114, 306, 482.
Photinia davidsoniae Rehder & Wilson, Cheng & Hwa 1118, Hwa 106.
Photinia franchetiana Diels, Cheng & Hwa 1001; Hwa 409, 523.
Photinia parvifolia Schneider, Cheng & Hwa 563, 831, 1094, Hwa 470.
Photinia sp. Cheng & Hwa 608, 799.
Prunus conradinae Koehne, Djou 126, Hwa 297, 358.
Prunus glyptocarpa Koehne, Hwa 60, 247, 263, 331, 349, 552.
Prunus grayana Maxim. Cheng & Hwa 703; Gressitt 2552; Hwa 285, 347, 550.
Prunus microbotrys Koehne, Cheng & Hwa 669, 737, 1149, Hwa 170, 338.
From Nepal extending eastward via China to Japan, there exists a group of species of evergreen Prunus characterized by subcoriaceous leaves that are eglandular beneath and by simple axillary racemose inflorescences. The names proposed for the species are Prunus wallichii Steudel (1841) for the Nepalese species, P. spinulosa Sieb. & Zucc. (1843) for the Japanese species, and P. microbotrys Koehne (1911) for the species from Central China. Kalkman (1965) renamed some of the Chinese specimens P. wallichii and others P. spinulosa (including P. microbotrys). However, it is thought that there are three species in this complex. Typical P. wallichii has large, thin leaves 12-13 cm. long and 5-7 cm. wide. The racemes are usually
[p. 78]
fasciculate, and the rachis and pedicels are glabrous. The basal portion of the ovary is pilose. Typical P. spinulosa from Japan has leaves that are smaller (6-9 cm. long, 1.5-3 cm. wide) and acute at the base. The racemes are always solitary, and the rachis and pedicels are strigose. The ovary is glabrous. Typical P. microbotrys from Central China has oblong or ovate-oblong leaves 6-9 cm. long and 2.5-3.5 cm. wide. The racemes are solitary, and the rachis and pedicels are pilose. The ovary is hairy at the base. Specimens from Central and West China are variable, especially in the size, shape, texture, and dentation of the leaves. These involve morphological characters that do not exist in the specimens from Japan, Nepal, and adjacent northern India. The name P. microbotrys, originally published for the taxon of Central and West China, should be maintained.
Prunus microbotrys Koehne var. obovata S. Y. Hu, var. nov.
Arbor, ramulis glabris; foliis glabris, oblongo-obovatis, 10-11 cm. longis, 4.5-5 cm. latis, basi acutis, apice caudato-acuminatis, acuminibus 1.2-1.4 cm. longis; racemis axillaribus, solitariis, 4-7 cm. longis, rachibus et pedicellis pilosis, calycibus glabris, petalis 2 mm. diametro.
Cheng & Hwa 735 (holotype, A).
Prunus mume (Sieb.) Sieb. & Zucc. Gressitt 2524 (sterile).
Prunus padus L. Gressitt 5248; Hwa 390.
Prunus persica (L.) Batsch, Djou 135.
Prunus pseudocerasus Lindley, Gressitt 2534.
Prunus salicina Lindley, Djou 109; Gressitt 2439: Hwa 256.
Prunus szechuanica Batalin, Hwa 248, 343.
Prunus venosa Koehne, Cheng & Hwa 816, Hwa 7.
Prunus wilsonii (Diels) Koehne, Gressitt 2570, Hwa 369.
Prunus wilsonii (Diels) Koehne var. leiobotrys Koehne, Cheng & Hwa 592.
Pyracantha crenato-serrata (Hance) Rehder, Cheng & Hwa 977, Djou 127, Gressitt 2426.
Pyrus calleryana Decaisne, Djou 106.
Pyrus pyrifolia (Burman f.) Nakai, Hwa 319.
Rosa helenae Rehder & Wilson, Djou 116.
Rosa roxburghii Tratt. Cheng & Hwa 956.
Rubus adenophorus Rolfe, Cheng & Hwa 637, 742.
Rubus ampelinus Focke, Cheng & Hwa 702.
Rubus cavaleriei Lévl. Cheng & Hwa 826; Gressitt 2432, 2519 (very closely allied to R. setchuensis Bur. & Franchet, but with leaf apex truncate).
Rubus ichangensis Hemsley & Kuntze, Cheng & Hwa 905.
Rubus malifolius Focke, Gressitt 2459.
Rubus pectinellus Maxim. Cheng & Hwa 591.
Sorbus aronioides Rehder, Cheng & Hwa 901, 1101.
Sorbus caloneura (Stapf) Rehder, Cheng & Hwa 671; Hwa 52, 311.
Sorbus folgneri (Schneider) Rehder, Cheng & Hwa 674, 1103; Hwa 58. 501, 553.
Sorbus wilsoniana Schneider, Cheng & Hwa 899, 1070.
Spiraea chinensis Maxim. Cheng & Hwa 945.
Spiraea japonica L. f. var. fortunei Rehder, Gressitt 2436.
Spiraea prunifolia Sieb. & Zucc. Hwa 398.
Stephanandra chinensis Hance, Cheng & Hwa 878.
Stranvaesia davidiana Decaisne var. undulata (Decaisne) Rehder & Wilson, Cheng & Hwa 900, 1034; Hwa 159, 325, 380.
[p. 79]
Leguminosae
AIbizia kalkora Prain, Cheng & Hwa 1054, Gressitt 2472; Hwa 37.
Amphicarpaea edgeworthii Bentham, Cheng & Hwa 832 (det. C. E. Wood, Jr.), 1144.
Cercis chinensis Bunge, Hwa 69 (sterile).
Cladrastis sinensis Hemsley, Gressitt 2461; Hwa 417.
Dalbergia dyeriana Prain, Cheng & Hwa 822.
Dalbergia mimosoides Franchet, Cheng & Hwa 804 (det. Y. T. Lee).
Desmodium podocarpum W. Cheng & Hwa 626, 820; Gressitt 2497.
Gleditsia macracantha Desf. Gressitt 2443; Hwa 45, 477.
Gymnocladus chinensis Baillon, Cheng & Hwa 1038; Hwa 478 (det. Y. T. Lee).
Indigofera amblyantha Craib, Gressitt 2514.
Indigofera pseudotinctoria Matsumura, Cheng & Hwa 1116 (det. Y. T. Lee).
Lespedeza cuneata G. Don, Cheng & Hwa 659, 727, Djou 128.
Lespedeza thunbergii (DC.) Nakai, Cheng & Hwa 895; Gressitt 2532.
Maackia chinensis Takeda, Cheng & Hwa 1146.
Ormosia puberula Cheng, Hwa 485 (isotype; det. W. C. Cheng).
Pueraria lobata (Willd.) Ohwi, Cheng & Hwa 731; Gressitt 2562.
Robinia pseudo-acacia L. Djou 139 (sterile; introduced).
Oxalidaceae
Oxalis griffithii Edgew. & Hooker f. Cheng & Hwa 787 (sterile; Dr. A. Lourteig consulted).
Rutaceae
Boenninghausenia albiflora (Hooker) Meissner, Cheng & Hwa 870.
Euodia fargesii Dode, Hwa 504.
Euodia meliaefolia Bentham, Hwa 57.
Euodia officinalis Dode, Cheng & Hwa 706, 1013; Gressitt 2421, 2442, 2475, 2540; Hwa 83.
Orixa japonica Thunb. Hwa 346.
Phellodendron chinense Schneider, Gressitt 2451; Hwa 13.
Toddalia asiatica (Lam.) Kurz, Cheng & Hwa 927, 942; Hwa 423.
Zanthoxylum dissitum Hemsley, Cheng & Hwa 601.
Zanthoxylum nitidum (Roxb.) DC. Hwa 337.
Zanthoxylum simulans Hance, Hwa 152.
Simaroubaceae
Ailanthus altissima (Miller) Swingle, Cheng & Hwa 720, 818, 1156; Djou 133; Gressitt 2554.
Meliaceae
Toona sinensis (A. Juss.) Roemer, Cheng & Hwa 1152; Hwa 151.
Polygalaceae
Polygala wattersii Hance, Hwa 5, 416.
Euphorbiaceae
Aleurites fordii Hemsley, Djou 136.
Croton tiglium L. Cheng & Hwa 1182.
[p. 80]
Glochidion pubescens (L.) Hutch. Cheng & Hwa 687, Djou 115, 124; Gressitt 2538.
Phyllanthus flexuosus (Sieb. & Zucc.) Mueller-Arg. Cheng & Hwa 648 (det. G. L. Webster).
Ricinus communis L. Cheng & Hwa 1183.
Sapium sebiferum (L.) Roxb. Cheng & Hwa 1181.
Daphniphyllaceae
Daphniphyllum glaucescens Bl. Cheng & Hwa 1030; Hwa 454.
Daphniphyllum longistylum Chien, Cheng & Hwa 660, 1072; Hwa 533, 534.
Coriariaceae
Coriaria sinica Maxim. Gressitt 2442, 2563; Hwa 238, 249, 250.
Buxaceae
Buxus microphylla Sieb. & Zucc. Cheng & Hwa 806.
Anacardiaceae
Choerospondias axillaris (Roxb.) Burtt & Hill, Cheng & Hwa 920, Hwa 447, 481, 498, 525, 555.
Rhus chinensis Miller, Djou 123; Gressitt 2470, 2533.
Rhus potaninii Maxim. Hwa 71.
Rhus trichocarpa Miq. Cheng & Hwa 744, 1104.
Rhus verniciflua Stokes, Cheng & Hwa 722; Gressitt 2473; Hwa 10, 149.
Aquifoliaceae
Ilex dasyphylla Merr. var. lichuanensis S. Y. Hu, var. nov.
A typo differt in ramulis petiolisque sparse pilosis; foliis subglabris, costa supra elevata; fructibus 6 mm. diametro, sepalis persistentibus glabris.
Cheng & Hwa 1055 (holotype, A).
In shape, size, and texture, the leaves of this variety are similar to those of Ilex dasyphylla Merr. var. dasyphylla, a variety of the mountainous area of northern Kwangtung and eastern Kwangsi, characterized by dense, ferrugineous, long hairs on the branchlets, leaves, and inflorescences. The fruit of var. lichuanensis is small, 4-5 mm. in diameter, and the persistent sepals are hirsute.
Ilex elmerrilliana S. Y. Hu, Cheng & Hwa 767, Hwa 487.
Ilex ficoidea Hemsley, Hwa 134 (sterile).
Ilex intermedia Loes. Hwa 132, 518, 521.
Ilex intermedia Loes. var. fangii (Rehder) S. Y. Hu, Cheng & Hwa 937.
Ilex macrocarpa Oliver, Hwa 97, 409, 520.
Ilex macropoda Miq. Cheng & Hwa 1111.
Ilex micrococca Maxim. Cheng & Hwa 1026.
Ilex pedunculosa Miq. Cheng & Hwa 1113.
Ilex pernyi Franchet, Cheng & Hwa 829, Hwa 4.
Ilex suaveolens (Lévl.) Loes. Cheng & Hwa 729, 1035; Hwa 41.
Ilex wilsonii Loes. Cheng & Hwa 775, 1139, Hwa 541.
Celastraceae
Celastrus gemmatus Loes. Djou 111.
Celastrus hypoleucus (Oliver) Warb. Cheng & Hwa 1062.
[p. 81]
Euonymus acanthocarpa Franchet, Cheng & Hwa 1037.
Euonymus acanthocarpa Franchet var. sutchuenensis Franchet ex Loes. Cheng & Hwa 1004.
Euonymus alata Regel, Hwa 112, 524.
Euonymus japonica Thunb. var. acuta Rehder, Cheng & Hwa 1121; Gressitt 2518, Hwa 102.
Euonymus myriantha Hemsley, Cheng & Hwa 949.
Euonymus sanguinea Loes. var. camptoneura Loes. Djou 122; Gressitt 2413; Hwa 82.
Aceraceae
Acer amplum Rehder, Cheng & Hwa 740; Hwa 8, 360.
Acer cordatum Pax, Hwa 428, 446, 540.
Acer davidii Franchet, Cheng & Hwa 714, 902, 1069, Hwa 288, 345.
Acer erianthum Schwerin, Cheng & Hwa 704, Gressitt 2565.
Acer faberi Hance, Cheng & Hwa 991; Hwa 428, 489.
Acer henryi Pax, Hwa 335 (a very young shoot), 472.
Acer longipes Franchet ex Rehder, Hwa 55.
Acer mono Maxim. Hwa 28.
Acer oliverianum Pax, Cheng & Hwa 862.
Acer sinense Pax, Cheng & Hwa 783; Hwa 139, 339.
Acer wilsonii Rehder, Cheng & Hwa 569, Hwa 348, 465.
Hippocastanaceae
Aesculus wilsonii Rehder, Cheng & Hwa 1134; Hwa 96 (sterile).
Sabiaceae
Meliosma beaniana Rehder & Wilson, Hwa 59, 67, 131, 545 (59 and 131 both sterile).
Meliosma pendens Rehder & Wilson, Cheng & Hwa 577, Gressitt 2460.
Meliosma sinensis Nakai, Cheng & Hwa 772.
Balsaminaceae
Impatiens lucorum Hooker f. Cheng & Hwa 642, 796, 838 (very young).
Staphyleaceae
Euscaphis japonica (Thunb.) Kanitz, Cheng & Hwa 768, Gressitt 2476, 2531; Hwa 565.
Tapiscia sinensis Oliver, Cheng & Hwa 564, Hwa 50, 538.
Rhamnaceae
Berchemia floribunda (Wall.) Brongn. Cheng & Hwa 621; Djou 119, Gressitt 2406, 2542.
Berchemia polyphylla Wall. ex Lawson, Cheng & Hwa 1046.
Hovenia trichocarpa Chun & Tsiang, Cheng & Hwa 561, 734, 1039; Hwa 168.
Rhamnus esquirolii Lévl. Cheng & Hwa 810.
Rhamnus paniculiflorus Schneider, Cheng & Hwa 1115.
Rhamnus rugulosus Hemsley, Cheng & Hwa 613, 1045.
Rhamnus utilis Decaisne, Cheng & Hwa 607, 629, 833, 909, 970, 971; Djou 131; Gressitt 2448, 2523; Hwa 68, 98, 512.
Sageretia henryi J. R. Drumm. Cheng & Hwa 864.
[p. 82]
Vitaceae
Ampelopsis chaffanjoni (Lévl.) Rehder, Cheng & Hwa 766, 827.
Parthenocissus heterophylla (Bl.) Merr. Cheng & Hwa 917.
Tetrastigma serrulatum Planchon, Cheng & Hwa 943.
Elaeocarpaceae
Elaeocarpus chinensis (Gardner & Champion) Hooker f. Cheng & Hwa 680; Hwa 434.
Sloanea hemsleyana (Ito) Rehder & Wilson, Cheng & Hwa 1097; Hwa 293.
Tiliaceae
Tilia tuan Szysz. Cheng & Hwa 741, 1059, 1114.
Tilia sp., Cheng & Hwa 1095.
Malvaceae
Hibiscus syriacus L. Cheng & Hwa 721; Hwa 99.
Actinidiaceae
Actinidia arguta Planchon ex Miq. Cheng & Hwa 1109.
Actinidia callosa Lindley, Cheng & Hwa 606.
Actinidia chinensis Planchon, Gressitt 2419, 2495.
Actinidia coriacea (Finet & Gagnep.) Dunn, Cheng & Hwa 567.
Theaceae
Camellia caudata Wall. Cheng & Hwa 620, 931.
Camellia cuspidata (Kochs) Veitch, Cheng & Hwa 609, 686, Gressitt 2471; Hwa 270.
Camellia japonica L. Hwa 255.
Camellia oleifera Abel, Cheng & Hwa 568, 1044, Gressitt 2450.
Camellia sinensis (L.) O. Kuntze, Cheng & Hwa 689, 1140, Gressitt 2490; Hwa 320.
Eurya nitida Korthals var. aurescens (Rehder & Wilson) Kobuski, Hwa 136, 273.
Schima sinensis (Hemsley & Wilson) Airy Shaw, Cheng & Hwa 890; Gressitt 2480.
Schima superba Gardner & Champion, Cheng & Hwa 1023.
Schima parviflora Cheng & H. T. Chang, Cheng & Hwa 953 (isotype; det. W. C. Cheng).
Ternstroemia nitida Merr. Hwa 455 (flower buds very young).
Guttiferae
Hypericum ascyron L. Gressitt 2431.
Hypericum attenuatum Choisy, Cheng & Hwa 819.
Hypericum hookerianum Wight & Arnott, Cheng & Hwa 593.
Hypericum patulum Thunb. Cheng & Hwa 624.
Hypericum petiolulatum Hooker f. & Thomson, Cheng & Hwa 736, 1092
Violaceae
Viola cf. confusa Champion ex Bentham, Cheng & Hwa 1087 (sterile).
Viola inconspicua Bl. Cheng & Hwa 965, 1091.
Viola triangulifolia W. Beck, Cheng & Hwa 1086.
[p. 83]
Flacourtiaceae
Carrierea calycina Franchet, Cheng & Hwa 928.
Idesia polycarpa Maxim. Cheng & Hwa 723, 979, Djou 118; Hwa 539.
Idesia polycarpa Maxim. var. vestita Diels, Cheng & Hwa 1060.
Poliothyrsis sinensis Oliver, Cheng & Hwa 1132; Hwa 511.
Stachyuraceae
Stachyurus himalaicus Hooker f. & Thomson, Cheng & Hwa 604, 988, 994.
Begoniaceae
Begonia evansiana Andr. Cheng & Hwa 590.
Begonia pedatifida Lévl. Cheng & Hwa 840.
Elaeagnaceae
Elaeagnus bockii Diels, Cheng & Hwa 800, Gressitt 2462.
Elaeagnus henryi Warb. Hwa 158.
Elaeagnus schnabeliana Hand.-Mazz. Cheng & Hwa 608.
Elaeagnus viridis Servettaz, Cheng & Hwa 809.
Lythraceae
Lagerstroemia indica L. Cheng & Hwa 765.
Alangiaceae
Alangium chinense (Lour.) Rehder, Cheng & Hwa 678, Gressitt 2492.
Alangium handelii Schnarf, Hwa 105 (closely related to A. chinense; recognized by hairy fruits and strigose branchlets and inflorescences).
Nyssaceae
Nyssa sinensis Oliver, Cheng & Hwa 611; Hwa 61, 432, 433.
Melastomataceae
Sarcopyramis nepalensis Wall. Cheng & Hwa 572, 730 (very small and young; more material needed for positive determination).
Onagraceae
Circaea erubescens Franchet & Sav. Cheng & Hwa 842.
Epilobium angustifolium L. Cheng & Hwa 712.
Araliaceae
Aralia decaisneana Hance, Cheng & Hwa 888.
Dendropanax chevalieri (Vig.) Merr. Cheng & Hwa 684; Hwa 156, 174 (both sterile).
Evodiopanax evodiifolius (Franchet) Nakai, Cheng & Hwa 900, 903, 980, Hwa 464.
Hedera nepalensis K. Koch var. sinensis (Tobler) Rehder, Cheng & Hwa 780, 1129, Hwa 129.
Kalopanax septemlobus (Thunb.) Koidzumi (Kalopanax pictus (Thunb.) Nakai), Cheng & Hwa 711; Hwa 76.
Nothopanax bodinieri (Lévl.) S. Y. Hu, comb. nov.
Aralia bodinieri Lévl. In Bull. Geogr. Bot. 24: 143. 1914.
Nothopanax delavayi sensu Rehder in Jour. Arnold Arb. 15: 115. 1934, pro parte.
[p. 84]
Rehder in 1934 interpreted Heptapleurum esquirolii Lévl. (Cavalerie 871, holotype), and Aralia bodinieri Lévl. (Cavalerie 2696, syntype) as synonyms of Nothopanax delavayi (Franchet) Harms. He was correct with the first epithet. Fragments and a photograph of the syntype of Aralia bodinieri Lévl. and a specimen with good fruiting material (Cheng & Hwa 969) show distinctive characteristics in leaves and inflorescences that do not appear in over 40 sheets of Yunnan specimens of Nothopanax delavayi examined. Aralia bodinieri (Cavalerie 2696) is a species of Nothopanax and is transferred above. Nothopanax bodinieri has unifoliolate, trifoliolate, and pentafoliolate leaves with the leaflets 2-4 times wider than those of N. delavayi. The fruiting peduncles of the umbels are half as long as those of N. delavayi. It is a distinctive species of northeastern Kweichow and the adjacent areas of Szechwan and Hupeh, while N. delavayi is endemic to western Yunnan.
Nothopanax davidii (Franchet) Harms, Cheng & Hwa 783; Gressitt 2525; Hwa 43.
Nothopanax rosthornii Harms, Cheng & Hwa 773; Gressitt 2556.
Schefflera delavayi (Franchet) Harms, Cheng & Hwa 978.
Schefflera venulosa (Wight & Arnott) Harms, Hwa 72.
Cornaceae
Aucuba chinensis Bentham f. angustifolia Rehder, Hwa 315.
Cornus controversa Hemsley, Cheng & Hwa 634; Hwa 458.
Cornus kousa Buerger var. chinensis Bean, Cheng & Hwa 570, 664, 908; Hwa 31, 107.
Cornus macrophylla Wall. Cheng & Hwa 632, 919, 1014; Djou 137, Gressitt 2428; Hwa 29, 91, 130.
Helwingia japonica (Thunb.) Dietrich, Cheng & Hwa 640, 930, 1093; Gressitt 2429.
Torricellia intermedia Harms, Cheng & Hwa 1145, Hwa 12, 421.
Umbelliferae
Daucus carota L. Djou 130; Gressitt 2520.
Clethraceae
Clethra fargesii Franchet, Cheng & Hwa 821, 1068, 1110.
Ericaceae
Enkianthus chinensis Franchet, Cheng & Hwa 1100.
Enkianthus serrulatus Schneider, Cheng & Hwa 616, Hwa 171, 275.
Lyonia ovalifolia (Wall.) Drude, Cheng & Hwa 746, 1053; Hwa 73.
Pieris formosa D. Don, Cheng & Hwa 802, 1041; Hwa 44, 243.
Rhododendron argyrophyllum Franchet, Hwa 66, 274.
Rhododendron caeruleum Lévl. Cheng & Hwa 891.
Rhododendron ciliicalyx Franchet, Hwa 277.
Rhododendron farrerae Tate ex Sweet, Cheng & Hwa 897, 1027, Hwa 377.
Rhododendron fortunei Lindley, Cheng & Hwa 896, Hwa 353.
Rhododendron lutescens Franchet, Hwa 278.
Rhododendron ovatum (Lindley) Planchon, Cheng & Hwa 1029, Hwa 298, 445.
Rhododendron oxyphyllum Franchet, Cheng & Hwa 682; Hwa 84.
Rhododendron simsii Planchon, Cheng & Hwa 879, Gressitt 2417; Hwa 453.
[p. 85]
Vaccinium bracteatum Thunb. Cheng & Hwa 990.
Vaccinium carlesii Dunn, Hwa 299.
Vaccinium hirtum Thunb. Cheng & Hwa 605, 675, 1135.
Vaccinium iteophyllum Hance, Cheng & Hwa 610.
Vaccinium japonicum Miq. Cheng & Hwa 685.
Vaccinium mandarinorum Diels, Cheng & Hwa 673, 898, 1056.
Myrsinaceae
Ardisia chinensis Bentham, Cheng & Hwa 1154.
Ardisia crenata (Thunb.) DC. Cheng & Hwa 1031.
Maesa hupehensis Rehder, Cheng & Hwa 922.
Myrsine cf. semiserrata Wall. Cheng & Hwa 1148 (leaves and inflorescences match well, but uniformly punctate lower leaf surface differs).
Primulaceae
Lysimachia fortunei Maxim. Cheng & Hwa 1085.
Ebenaceae
Diospyros lotus L. Cheng & Hwa 710; Hwa 30.
Styracaceae
Alniphyllum megaphyllum Hemsley, Cheng & Hwa 984, 1008, Hwa 490.
Pterostyrax hispidus Sieb. & Zucc. Cheng & Hwa 1107, Hwa 491, 566.
Styrax bodinieri Lévl. Cheng & Hwa 559, 662, 975; Gressitt 241, 2573; Hwa 16, 427 (young inflorescences of some specimens infested by insect galls).
Styrax suberifolius Hooker f. & Arn. Cheng & Hwa 677, 918; Hwa 121.
Symplocaceae
Symplocos anomala Brand, Cheng & Hwa 770, 1064; Hwa 147.
Symplocos botryantha Franchet, Cheng & Hwa 690, 868, 904; Hwa 74, 140, 316, 373, 382.
Symplocos caudata Wall. Cheng & Hwa 603, 1061; Hwa 244, 310, 314.
Symplocos discolor Brand, Hwa 280, 544.
Symplocos lancifolia Sieb. & Zucc. Cheng & Hwa 691, 984, Djou 107, Hwa 172, 309.
Symplocos stellaris Brand, Cheng & Hwa,1112; Hwa 425.
Oleaceae
Fraxinus chinensis Roxb. Cheng & Hwa 807; Djou 101 (sterile); Hwa 402.
Fraxinus punctata S. Y. Hu, sp. nov.
Arbor vel frutex, glabra, ramulis robustis, hornotinis 4-5 mm. diametro, lenticellis albidis numerosis notatis; foliis trifoliolatis, petiolis 3.5-5 cm. longis, canaliculatis, basi incrassatis, foliolis petiolulis medianis 2-3 cm. longis, lateralibus 6-8 mm. longis, laminis subcoriaceis, ovatis, medianis 10-13 cm. longis, 6-7 cm. latis, lateralibus 8.5-12 cm. longis, 4-5 cm. latis, basi acutis, apice acuminatis, acumine 1-1.5 cm. longis, subtus glabris et punctatis, margine crenulato-serratis, costa supra impressa, subtus elevata et glabra, nervis lateralibus utrinque 15-17, subtus prominentibus, nervis secondariis bene reticulatis; infructescentiis terminalibus; samaris lineari-spathulatis, 2.5-3 cm. longis, 5 mm. latis, apice retusis; calycibus persis-
[p. 86]
tentibus patelliformibus, lobis deltoideis, apice subulato-acuminatis vel acutis.
Cheng & Hwa 946 (holotype, A).
The trifoliolate, subcoriaceous leaves and the linear-spathulate samaras of this species suggest a close relationship to Fraxinus trifoliolata W. W. Smith of the drainage basin of the Yangtze River in northwestern Yunnan. The lower leaf surface of the Yunnan species is densely white-hirsute, and the persistent calyx of the fruit is subcampanulate with very shallow erose lobes. The ovary of this species is covered by peltate glandular scales that are persistent in fruit and become rather conspicuous on the basal one-third of the samara. In contrast, the lower leaf surface of the species in the metasequoia area is glabrous and punctate, and its persistent calyx is patelliform. and has well-defined lobes that are subulate-acuminate or acute at the apex. The glandular scales on the fruit are very sparse and obscure.
Jasminum lanceolarium Roxb. Cheng & Hwa 989.
Ligustrum lucidum Aiton, Cheng & Hwa 1157, Hwa 68.
Ligustrum quihoui Carr. Cheng & Hwa 713.
Ligustrum sinense Lour. Cheng & Hwa 1077, Hwa 115.
Osmanthus cooperi Hemsley, Hwa 430.
Osmanthus fragrans Lour. Cheng & Hwa 724, Hwa 272, 303.
Loganiaceae
Buddleia davidii Franchet, Cheng & Hwa 811; Gressitt 2501; 2527.
Gentianaceae
Swertia bimaculata Clarke, Cheng & Hwa 955.
Apocynaceae
Trachelospermum jasminoides (Lindley) Lemaire, Hwa 261.
Asclepiadaceae
Cynanchum auriculatum Royle, Cheng & Hwa 638A; Djou 114, 132.
Convolvulaceae
Cuscuta chinensis Lam. Cheng & Hwa 860.
Verbenaceae
Callicarpa bodinieri Lévl. var. giraldii (Rehder) Rehder, Cheng & Hwa 1124, Gressitt 2526.
Callicarpa rubella Lindley var. hemsleyana Diels, Cheng & Hwa 1042.
Clerodendron bungei Steudel, Cheng & Hwa 631, 914; Gressitt 2454, 2553.
Clerodendron mandarinorum Diels, Cheng & Hwa 769, 894, Hwa 40.
Clerodendron trichotomum Thunb. Cheng & Hwa 913.
Premna puberula Pampan. Cheng & Hwa 560, Gressitt 2427, 2483, 2558.
Labiatae
Mosla chinensis Maxim. Cheng & Hwa 788, 1096.
Mosla dianthera (Buch.-Ham.) Maxim. Cheng & Hwa 789.
Perilla frutescens (L.) Britton, Gressitt 2567.
Pogostemon glaber Bentham, Cheng & Hwa 835.
[p. 87]
Prunella vulgaris L. Cheng & Hwa 793.
Teucrium bidentatum Hemsley, Cheng & Hwa 1032.
Solanaceae
Solanum lyratum Thunb. Cheng & Hwa 1177.
Scrophulariaceae
Botryopleuron nervosum (Hemsley) Hemsley, Cheng & Hwa 1015.
Paulownia fargesii Franchet, Hwa 95, 462.
Phtheirospermum chinense Bunge ex Fischer & Meyer, Cheng & Hwa 1088.
Bignoniaceae
Catalpa fargesii Bur. Cheng & Hwa 855, 935; Gressitt 2418; Hwa 510.
Gesneriaceae
Didissandra cf. sesquifolia C. B. Clarke, Cheng & Hwa 718, 841 (specimens resemble D. sesquifolia of northern India in having pubescent leaves and short-pedicellate flowers, but differ in having leaf margins subentire, not subduplo-serrate).
Didymocarpus hwaianus S. Y. Hu, sp. nov.
Herbae succulentae, perennes, glabres; internodis caulium 4-7 cm. longis, 3 mm. crassis; foliis oppositis, sessilibus, integris, ellipticis, 12-15 cm. longis, 4.5-5 cm. latis, supra glabris vel leviter hirsutis, subtus glabris, basi cuneatis, decurrentibus et connato-perfoliatis, apice acuminatis; inflorescentiis axillaribus, pedunculis 5 mm. longis, ramis subcymosis, axibus secondariis 6-10 mm. longis, pedicellis 3-4 mm. longis; sepalis foliaceis, fere ad basim libris, obovato-oblongis, 8-9 mm. longis, 5-6 mm. latis, glabris, apice rotundatis; corollis et staminibus ignotis; capsulis juvenilibus compresso-cornutis, 2.5 cm. longis, 3 mm. latis; annulis interruptis; stylis 1 cm. longis, stigmatibus inaequaliter bilobatis.
Cheng & Hwa 923 (holotype, A).
The foliaceous sepals free almost to the base, the short axillary inflorescences and the laterally compressed, curved, cornute young capsules of this species suggest a close relationship with Didymocarpus bicornutus (Hayata) S. Y. Hu,[5] comb. nov., from Taiwan, which has petiolate leaves. Another closely related species is D. tibeticus Franchet, which differs in having villose, lanceolate sepals and petiolate leaves.
Lysionotus pauciflora Maxim. Cheng & Hwa 1003.
Acanthaceae
Hemigraphis cuneata S. Y. Hu, sp. nov.
Herba prostrata, caulibus quadrangulatis, glabrescentibus; foliis parvis, rhomboideis, 1-2.5 cm. longis, 0.7-2 cm. latis, basi cuneatis, apice acutis, margine remote serratis, utrinque glabrescentibus, petiolis 5-15 mm. longis, glabris; floribus solitariis, in axillis foliorum sessilibus, bracteis foliaceis, obovato-cuneatis, 3 mm. longis, 2 mm. latis, apice obtusis; calycibus glabris, fere ad basim partitis, laciniis linearibus, 1.5 cm. longis, 1.5 mm. latis
[5] Didymocarpus bicornutus (Hayata) S. Y, Hu, comb. nov.
Chirita bicornuta Hayata, Ic. Pl. Formos. 3: 154. 1913.
[p. 88]
apice obtusis; capsulis oblongis, ad basim constrictis, 12-13 mm. longis, 2 mm. diametro, glabris; seminibus plano-compressis, suborbicularibus, 1.5-2 mm. diametro, atro-fuscis, papillosis.
Cheng & Hwa 887 (holotype, A).
This species is related to Hemigraphis procumbens (Lour.) Merr., from which it is readily distinguished by its glabrous leaves, sepals, and capsules. The leaves of H. procumbens are hirsute and the sepals hispid-strigose; the capsules are hairy at the apex.
Peristrophe roxburghiana (Schultes) Bremek. Cheng & Hwa 715.
Plantaginaceae
Plantago major L. Cheng & Hwa 717.
Rubiaceae
Emmenopteryx henryi Oliver, Cheng & Hwa 1173; Hwa 166.
Galium bungei Steudel, Cheng & Hwa 784, 1143.
Hedyotis lindleyana Hooker var. glabra Hara, Cheng & Hwa 790.
Lasianthus hartii Franchet, Cheng & Hwa 622.
Paederia chinensis Hance, Cheng & Hwa 926, 973, 996.
Paederia scandens (Lour.) Merr. Cheng & Hwa 628, 824.
Serissa foetida Commerson, Cheng & Hwa 983.
Caprifoliaceae
Abelia myrtilloides Rehder, Cheng & Hwa 861, 1133.
Abelia uniflora R. Br. Hwa 452.
Lonicera henryi Hemsley, Cheng & Hwa 676, 1036, 1083.
Lonicera macranthoides Hand.-Mazz. Cheng & Hwa 882.
Sambucus racemosa L. Hwa 11, 372.
Viburnum ichangense (Hemsley) Rehder, Cheng & Hwa 814, 1105, Gressitt 2559.
Viburnum ovatifolium Rehder, Cheng & Hwa 649, 992; Hwa 46.
Viburnum propinquum Hemsley, Cheng & Hwa 867; Hwa 265, 537.
Viburnum propinquum Hemsley var. mairei W. W. Sm. Cheng & Hwa 860, 1122.
Viburnum rhytidophyllum Hemsley, Hwa 359.
Viburnum rosthornii Graebner var. xerocarpa Loes. Cheng & Hwa 892; Hwa 47, 535.
Viburnum setigerum Hance, Chang & Hwa 803, 993, 1071; Djou 117; Gressitt 2434, 2521.
Viburnum sympodiale Graebner, Hwa 341 or 355 (number not clear).
Viburnum ternatum Rehder, Cheng & Hwa 589, 732, 1019; Gressitt 2474; Hwa 56.
Viburnum theiferum Rehder, Hwa 473.
Viburnum tomentosum Thunb. Hwa 282, 426.
Viburnum tsangii Rehder, Cheng & Hwa 883, 1161.
Weigela japonica Thunb. var. sinica (Rehder) Bailey, Cheng & Hwa 573; Hwa 471.
Valerianaceae
Patrinia villosa (Thunb.) Juss. ex DC. Cheng & Hwa 964.
[p. 89]
Cucurbitaceae
Gymnostemma pentaphyllum Makino, Cheng & Hwa 925, 1014.
Campanulaceae
Campanumoea javanica Bl. Cheng & Hwa 625; Gressitt 2424.
Lobelia davidii Franchet, Cheng & Hwa 643 (very similar to L. fangiana (E. Wimmer) S. Y. Hu, comb. nov., [6] from which it may be distinguished by the subglabrous ovary and hairy anthers).
Lobelia radicans Thunb. Cheng & Hwa 578.
Pratia nummularia (Lam.) A. Br. & Ascherson (Lobelia angulata Forster), Cheng & Hwa 599.
Compositae
Adenocaulon adhaerescens Maxim. Cheng & Hwa 719.
Ainsliaea fulvipes Jeffrey, Cheng & Hwa 786,
Anaphalis margaritacea (L.) Bentham & Hooker f. Cheng & Hwa 762.
Carpesium abrotanoides L. Cheng & Hwa 679.
Eupatorium chinense L. Cheng & Hwa 967.
Eupatorium japonicum Thunb. Cheng & Hwa 837.
Ixeris gracilis (DC.) Stebbins, Cheng & Hwa 1084.
Leontopodium sinense Hemsley, Gressitt 2515.
Ligularia duciformis (C. Winkler) Hand.-Mazz. Cheng & Hwa 850.
Monocotyledoneae
Alismataceae
Sagittaria pygmaea Miq. Cheng & Hwa 761.
Sagittaria sagittifolia L. subsp. sagittata (Thunb.) Rataj. Cheng & Hwa 760.
Hydrocharitaceae
Ottelia alismoides (L.) Pers. Cheng & Hwa 847 (material too young to show species characteristics).
Gramineae
Chimonobambus sp., Hwa 495, 496 (sterile; det. T. R. Soderstrom).
Miscanthus sinensis Andersson, Gressitt 2555.
Phyllostachys sp., Gressitt 2410 (sterile; det. T. R. Soderstrom).
Sinarudinaria sp., Gressitt 2494 (det. T. R. Soderstrom); Hwa 494.
[6] Lobelia fangiana (E. Wimmer) S. Y. Hu, comb. nov.
Pratia fangiana E. Wimmer, Repert. Sp. Nov. 38: 3. 1935.
Lobelia omiensis E. Wimmer, Ann. Nat. Mus. Wien 56: 366. 1948.
SZECHWAN: Mt. Omei, W. P. Fang 2853 (holotype, NY; isotype, GH), E. H. Wilson 5034 (isotype of L. omiensis, GH), H. C. Chow 7927, 8106 (A), S. C. Sum & K. Chang 1139, 1227 (A), L. Y. Tai 87 (A), F. T. Wang 23376 (A), K. N. Yin 127 (A). Wimmer's material of Pratia fangiana is in fruit, and his specimen of Lobelia omiensis is in flower. More recent collections from the same area give a fuller picture of morphological variation of the species, and the gaps caused by age differences in Wimmer's samples are bridged. The species is characterized by hirsute inflorescences, pubescent ovaries, and glabrous anthers. A transfer of the earlier specific epithet from Pratia to Lobelia is necessitated.
[p. 90]
Cyperaceae
Bulbostylis capillaris (L.) Clarke, Cheng & Hwa 1089.
Carex brunnea Thunb. Cheng & Hwa 791 (det. T. Koyama).
Cyperus orthostachys Franchet & Sav. Cheng & Hwa 753 (det. T. Koyama).
Eleocharis congesta D. Don subsp. japonica (Miq.) Koyama, Cheng & Hwa 758 (det. T. Koyama).
Kyllinga brevifolia Rottb. Cheng & Hwa 596, 757.
Pycreus flavescens (Poiret) Koyama, Cheng & Hwa 754 (det. T. Koyama).
Pycreus sanguinolentus Vahl, Cheng & Hwa 755 (det. T. Koyama).
Scirpus wallichii Roxb. Cheng & Hwa 756 (det. T. Koyama).
Eriocaulaceae
Eriocaulon buergerianum Koern. Cheng & Hwa 759.
Commelinaceae
Commelina communis L. Cheng & Hwa 785, 957.
Pontederiaceae
Monochoria vaginalis (Burman f.) Presl, Cheng & Hwa 763, 848.
Juncaceae
Juncus setchuensis Buchenau, Cheng & Hwa 644.
Liliaceae
Cardiocrinum giganteum (Wall.) Makino, Cheng & Hwa 849.
Disporum cantoniense (Lour.) Merr. Cheng & Hwa 941.
Lilium leucanthum Baker var. chloraster (Baker) Wilson, Gressitt 2513.
Paris chinensis Franchet, Cheng & Hwa 1033.
Protolirion sinii Krause, Cheng & Hwa 623.
Reineckea carnea Kunth, Cheng & Hwa 597.
Smilax lancifolia Roxb. Cheng & Hwa 982.
Smilax megalantha A. DC. Cheng & Hwa 633; Hwa 350.
Smilax myrtillus DC. Cheng & Hwa 929.
Smilax riparia DC. Cheng & Hwa 972, 974, 1155.
Iridaceae
Belamcanda chinensis (L.) DC. Gressitt 2416.
Iris japonica Thunb. Cheng & Hwa 652.
Dioscoreaceae
Dioscorea cirrhosa Lour. Cheng & Hwa 1018.
Dioscorea kamaonensis Kunth var. fargesii (Franchet) Prain & Burkill, Cheng & Hwa 638, 656, 836, 1016 (det. B. G. Schubert).
Orchidaceae
Cymbidium ensifolium Sw. Cheng & Hwa 1006.
Cypripedium japonicum Thunb. Cheng & Hwa 1162.
Pleione henryi Rolfe, Cheng & Hwa 911.
[p. 91]
ACKNOWLEDGMENTS
Grateful acknowledgment is due all my friends who have helped with the identification of specimens and the preparation of the manuscript. I should like to take this opportunity to express my special appreciation to Drs. L. M. Perry, B. G. Schubert, S. A. Spongberg, P. F. Stevens, R. M. Tryon, and C. E. Wood, Jr., for numerous helpful suggestions. Particular thanks are also extended to R. A. Howard for the opportunity to undertake this project. I should also like to acknowledge the careful work of Karen Stoutsenberger, who prepared the final versions of the maps that illustrate this article.
REFERENCES
ANDREWS, H. N., JR. 1961. Studies in paleobotany. xii + 487 pp. Wiley, New York.
Anonymous. 1948. Living fossil. Jour. N. Y. Bot. Gard. 49: 151.
---. 1949. Rand McNally cosmopolitan world atlas. xvi + 335 pp. Rand McNally & Co., New York.
---. 1972-1976. Iconographia Cormophytorum Sinicorum. Vol. 1. Bryophyta, Pteridophyta, Gymnospermae, Angiospermae (Casuarinaceae-Hermanniaceae). vi + 1157 pp. 1972. Vol. 11. Papaveraceae-Cornaceae. iv + 1312 pp. 1972. Vol. III. Diapensiaceae-Solanaceae. iii + 1083 pp. 1974. Vol. IV. Scrophulariaceae-Compositae. iii + 932 pp. 1975. Vol. V. Monocotyledoneae. iii + 1146 pp. 1976. Science Press, Peking.
BOUGHEY, A. S. 1965. Comparisons between the montane forest floras of N. America, Africa and Asia. Webbia 19(2): 506-517.
CHANEY, R. W. 1948. The bearing of the living metasequoia on problems of Tertiary paleobotany. Proc. Natl. Acad. Sci. U. S. A. 34: 503-515.
CHENG, W. C., L. K. Fu, et al. 1978. Flora Reip. Pop. Sin. 7: 34-54.
CHU, K. L., & W. S. COOPER. 1950. An ecological reconnaissance in the native home of Metasequoia glyptostroboides. Ecology 31: 260-278.
FLINT, R. F. 1957. Glacial and Pleistocene geology. xiii + 553 pp. Wiley, New York.
FLORIN, R. 1952. On metasequoia, living and fossil. Bot. Not. 1952: 1-29.
---. 1963. The distribution of conifer and taxad genera in time and space. Acta Horti Berg. 20: 121-312. [Additions and corrections. Ibid. 319-326. 1966.]
FLOUS, F. 1936. Espèces nouvelles de Keteleeria. Bull. Soc. Hist. Nat. Toulouse 69: 399-408.
FULLING, E. H. 1976. Metasequoia - fossil and living. Bot. Rev. 42: 215-315. [Additions. Ibid. 43: 281-284. 1977.]
GOOD, R. 1974. Geography of flowering plants. ed. 4. 557 pp. Longman, London.
GRESSITT, J. L. 1953. The California Academy - Lingnan dawn-redwood expedition. Proc. Cal. Acad. Sci. 28: 25-58.
HARA, H. 1966. Taxonomic comparison between corresponding taxa of
[p. 92]
Spermatophyta in eastern Himalaya and Japan. Pp. 627-657 in H. HARA, comp., The flora of eastern Himalaya. x + 744 pp. University of Tokyo Press, Tokyo.
Hu, H. H. How Metasequoia, the "living fossil," was discovered in China. Jour. N. Y. Bot. Gard. 49: 201-207.
--- & W. C. CHENG. 1948. On the new family Metasequoiaceae and on Metasequoia glyptostroboides, a living species of the genus Metasequoia found in Szechuan and Hupeh. Bull. Fan Mem. Inst. Biol. II. 1: 153-166.
KALKMAN, C. 1965. The Old World species of Prunus subg. Laurocerasus including those formerly referred to Pygeum. Blumea 13: 1-115.
LEE, J. S. 1939. The geology of China. 527 pp. Thomas Murby & Co., London.
LI, H. L. 1971. Floristic relationship between eastern Asia and eastern North America. Trans. Am. Philos. Soc. 42: 371-429.
MATHEWS, R. H. 1931. A Chinese-English dictionary. xxiv + 1226 pp. Presbyterian Mission Press, Shanghai.
MERRILL, E. D. 1948. Metasequoia, another "living fossil." Arnoldia 8: 4-8.
MIKI, S. 1941. On the change of flora in eastern Asia since Tertiary Period. 1. The clay or lignite bed flora in Japan with special reference to the Pinus trifolia beds in Central Hondo. Jap. Jour. Bot. 11: 237-303.
--- 1953. On Metasequoia, fossil and living. 141 pp. Nippon Kôbutsu Shumi-no, Kyoto.
--- 1954. The occurrence of the remains of Taiwania and Palaeotsuga (n. subg.) from Pliocene beds in Japan. Proc. Japan Acad. 30: 796-981.
NAIRN, A. E. M., ed. 1961. Descriptive palaeoclimatology. xi + 380 pp. Interscience Publishers, Inc., New York.
OHWI, J. Flora of Japan. (English edition; F. G. MEYER & E. H. WALKER, eds.) ix + 1067 pp. Smithsonian Institution, Washington, D. C.
ORR, M. Y. 1933. A new Chinese silver fir. Notes Bot. Gard. Edinburgh 18: 1-5.
---, 1933a. Plantae Chinenses forrestianae: Coniferae. Ibid. 119-158.
REHDER, A. 1940. Manual of cultivated trees and shrubs. ed. 2. xxx + 996 pp. Macmillan, New York.
---. 1949. Bibliography of cultivated trees and shrubs. xl + 825 pp. Arnold Arboretum, Jamaica Plain, Massachusetts.
--- & E. H. WILSON. 1914. Taxaceae and Pinaceae. In: C. S. SARGENT, ed., Pl. Wils. 2: 3-62.
SMALL, J. K. 1913. Flora of the southeastern United States. ed. 2. 1394 pp. "Published by the author," New York.
WANG, Z., & P. Y. Fu. 1974. Plantae novae Salicacearum Tibeticarum. Acta Phytotax. Sinica 12: 191-213.
WILSON, E. H. 1926. The taxads and conifers of Yunnan. Jour. Arnold Arb. 7: 37-68.
WOLFE, J. A. 1969. Neogene floristic and vegetational history of the Pacific Northwest. Madrono 20: 83-110.
--- 1978. A paleobotanical interpretation of Tertiary climates in the Northern Hemisphere. Am. Sci. 66: 694-703.
WOOD, C. E., JR. 1971. Some floristic relationships between the Southern Appalachians and western North America. Pp. 331-404 in P. C. HOLT, ed., The distributional history of the biota of the Southern Appalachians.
[p. 93]
Part II. Flora. Research Div. Monogr. 2. Virginia Polytech. Inst. State Univ. Blacksburg.
---. 1972. Morphology and phytogeography: the classical approach to the study of disjunctions. Ann. Missouri Bot. Gard. 59: 107-124.
ARNOLD ARBORETUM
22 DIVINITY AVENUE
CAMBRIDGE, MASSACHUSETTS 02138
[p. 94]
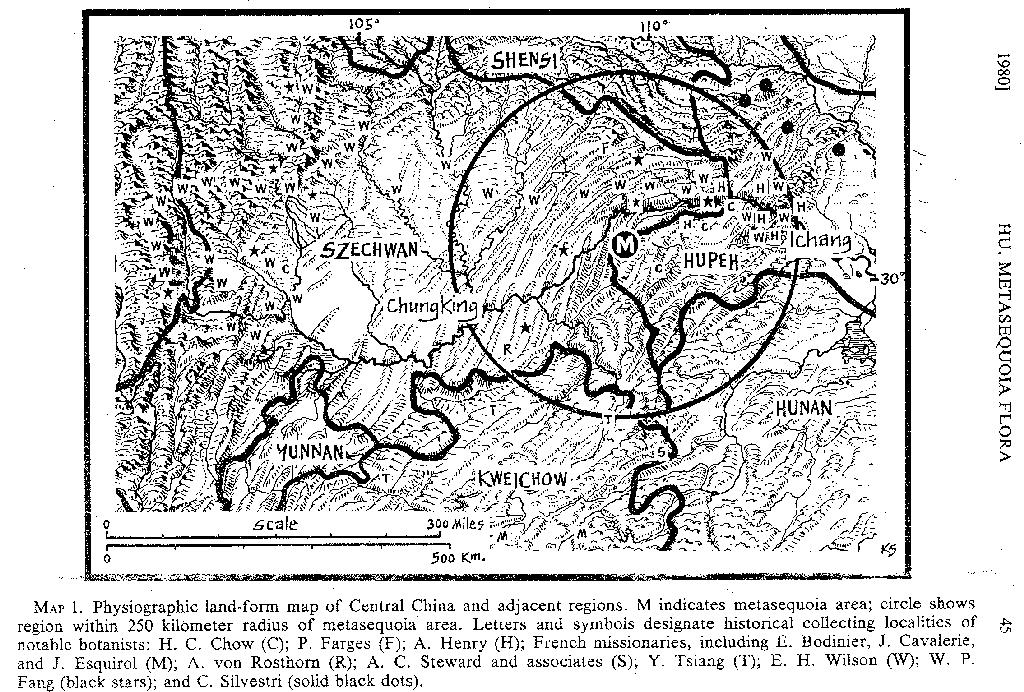{kind=link}
{kind=link}
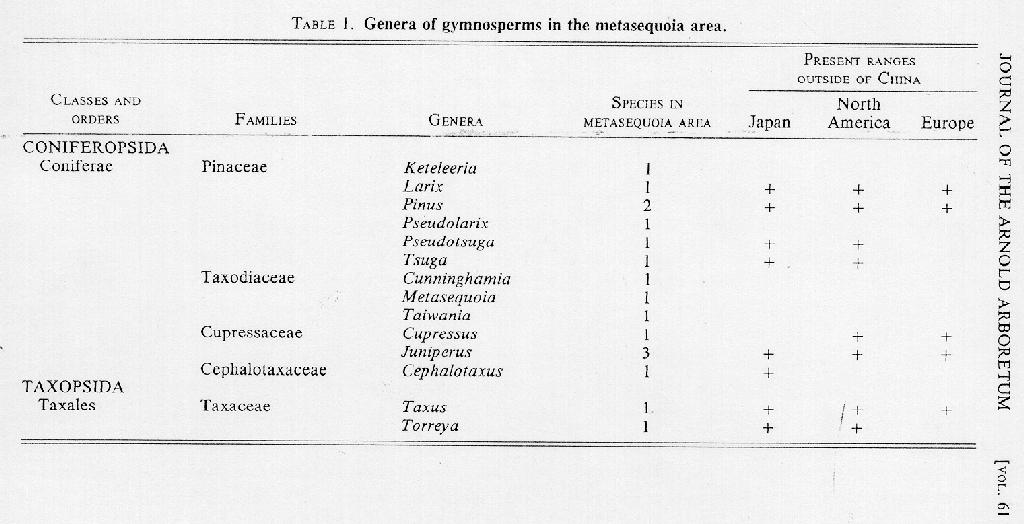{kind=link}
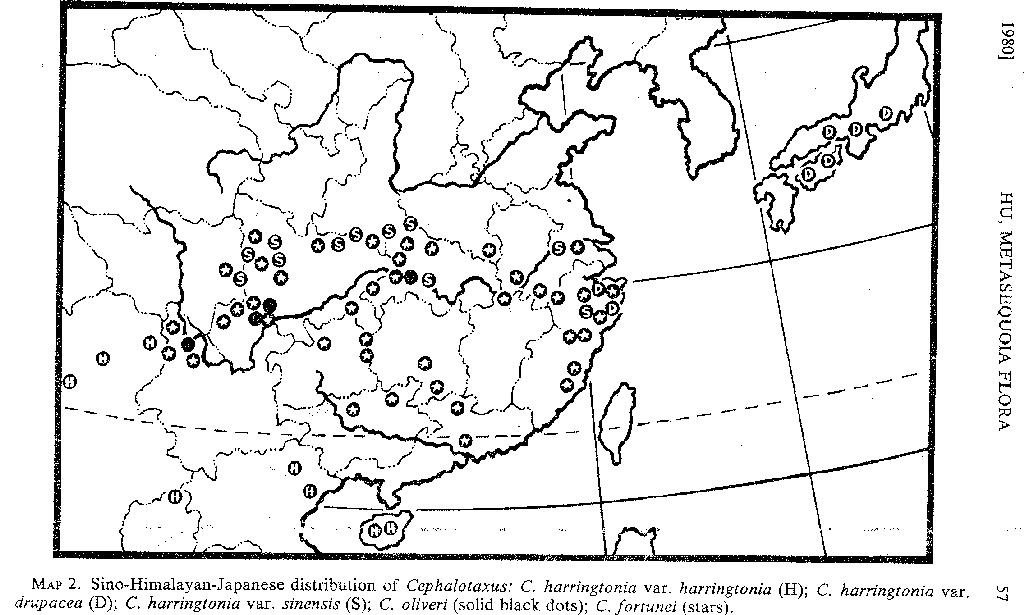{kind=link}
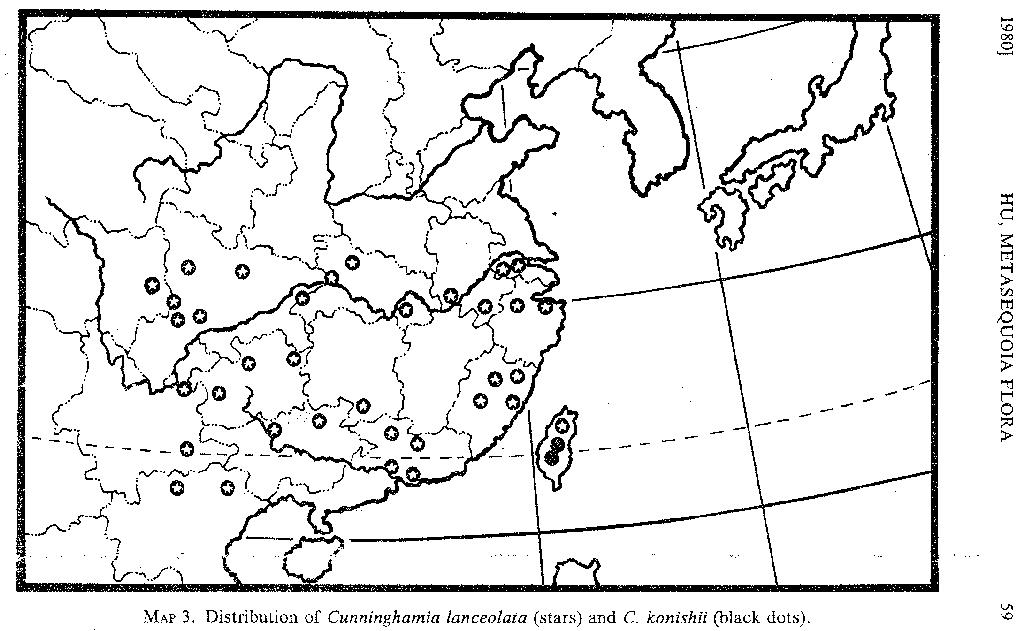{kind=link}
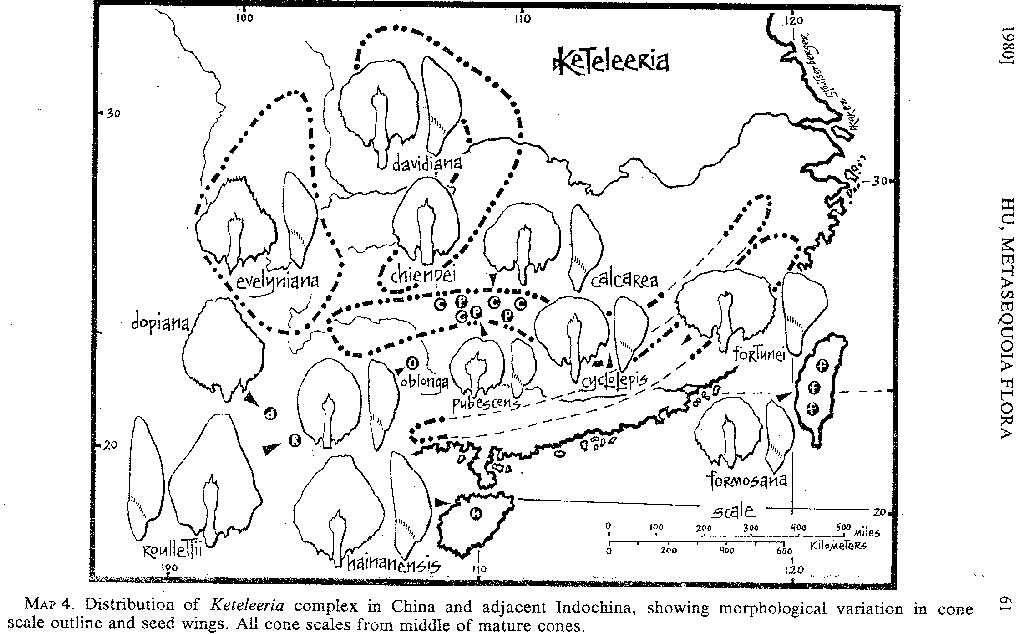{kind=link}
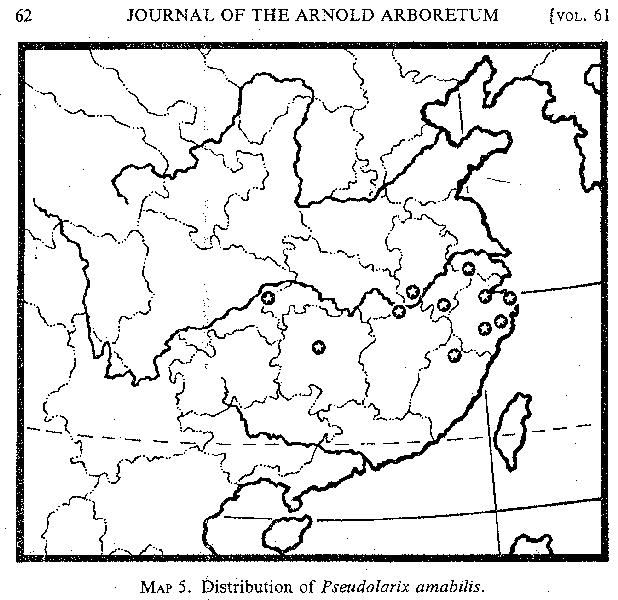{kind=link}
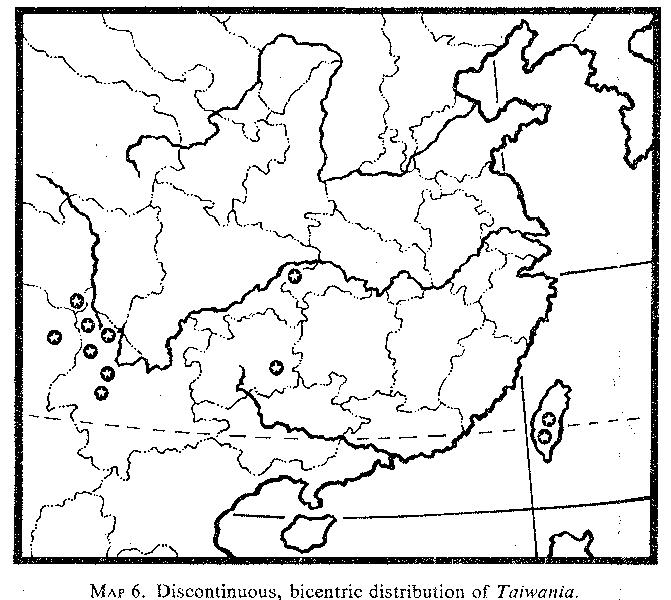{kind=link}
{kind=link}
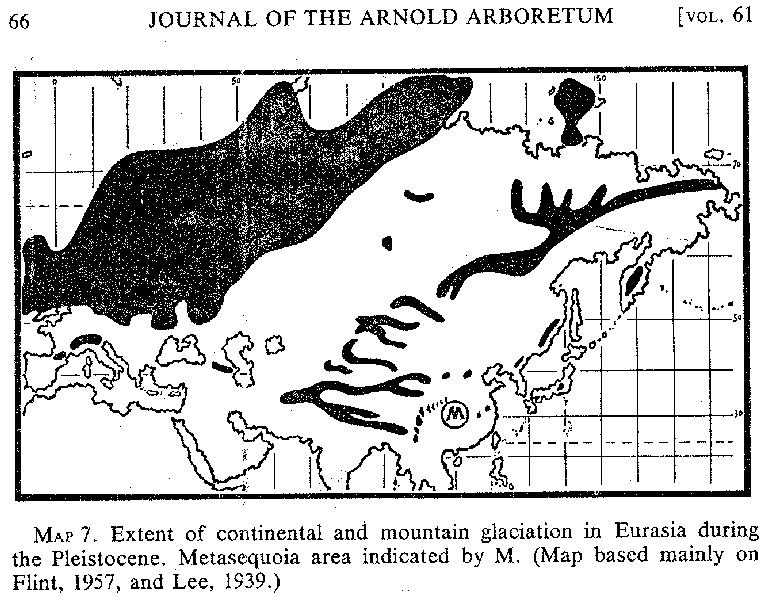{kind=link}
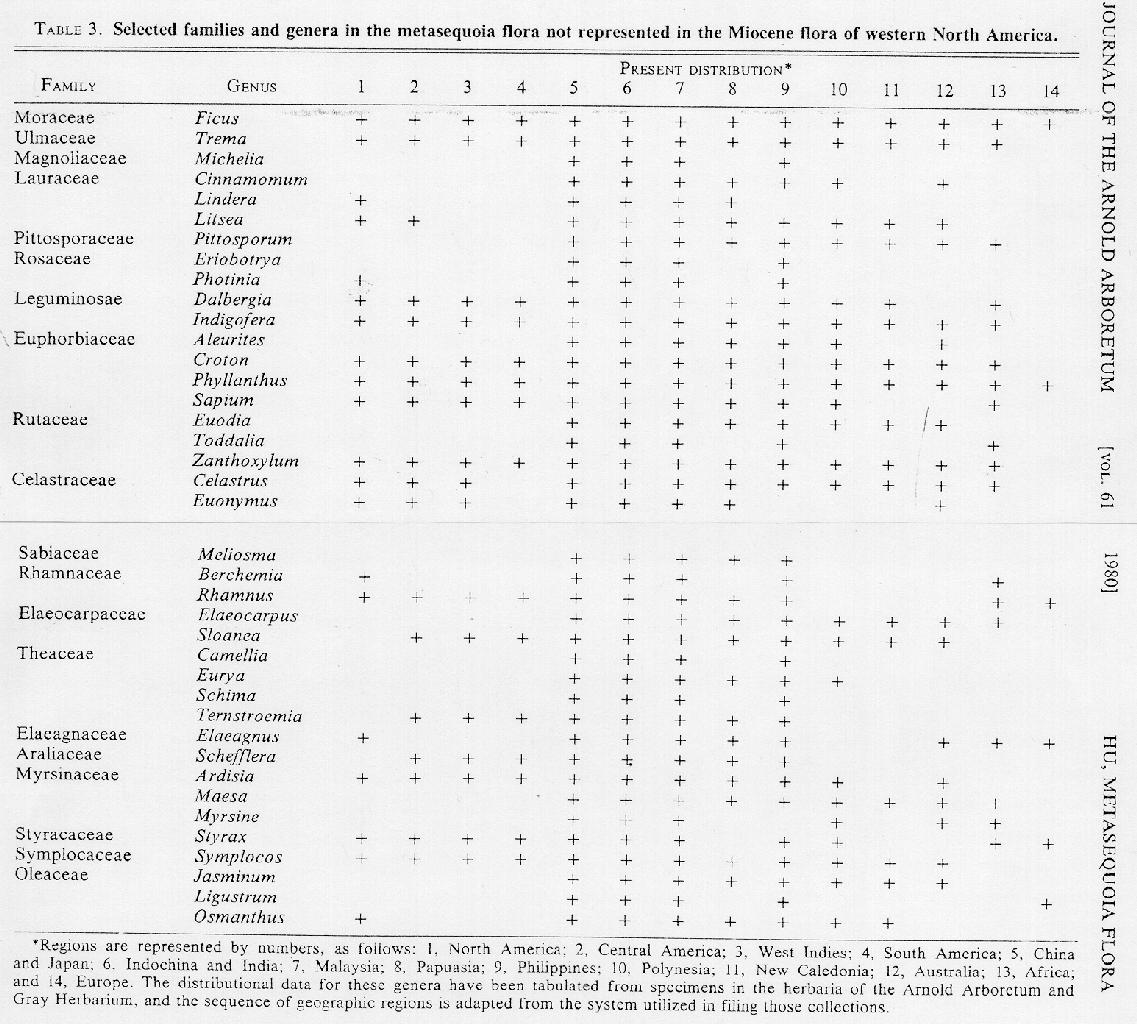{kind=link}