Ann. Missouri Bot. Gard. 85(4): 554-564. 1998.
Hong De-yuan2 , Pan Kai-yu2, and Yu Hong 2
Abstract
The Paeonia delavayi complex, a group of woody peonies endemic to China, is problematic because of the lack of agreement among the taxonomic treatments of the group. Some taxonomists recognize one species with three infraspecific taxa, while others accept only two species, and yet others accept three species with two infraspecific taxa. The present paper is based on extensive field observations, population sampling, and analysis of certain characters. The results show that the complex is extremely variable both within and between populations in the number, length, and width of leaf segments and in number, size, and color of all the floral parts. This variation is continuous, and there is no correlation between the characters. Only one species, P. delavayi, is recognized, without infraspecific taxa. Paeonia lutea, P. potaninii, and their infraspecific names are reduced to synonymy of P. delavayi. Vegetative reproduction by stolons is predominant in this variable species, and the roots are thickened fusiformly. These two features probably facilitate adaptation to open and somewhat dry habitats, and allow for rapid colonization whenever plants become established. Although P. delavayi has been listed as endangered, it is unlikely to become extinct if wanton digging is controlled.
The Paeonia delavayi (Paeoniaceae) complex comprises a group of woody peonies endemic to southwestern China. It and other shrubs of the genus Paeonia belong to section Moutan DC., while the herbaceous members belong to section Paeonia and section Onaepia Lindl. Members of section Moutan are readily distinguished by the following key:
1a. Flowers solitary and terminal, erect.
2a. Carpels nearly always 5, tomentose, entirely enveloped by floral disk at anthesis...P. suffruticosa complex
2b. Carpels 2--4(or 5), glabrous, enveloped by floral disk only in the lower half.....P. decomposita Hand.-Mazz.
1b. Flowers usually 2 or 3, both terminal and axillary, _ pendent.
3a. Carpels nearly always solitary; petals, filaments, and stigmas always pure yellow; plants 1.5--3.5 m tall; follicles 4.7--7 ´ 2--3.3 cm...P. ludlowii (Stern & G. Taylor) D. Y. Hong
3b. Carpels mostly 2--5(--7); petals, filaments, and stigmas variable in color; plants _ 2 m tall; follicles _ 4 ´ 1.5 cm....P. delavayi complex
Four species and numerous infraspecific taxa have been allied with Paeonia delavayi. The dark red-flowered P. delavayi Franch. and the yellow-flowered P. lutea Delavay ex Franch. were both based on collections from Northwest Yunnan [Lijiang (Likiang) and Eryuan, respectively] and were described on the same page (Franchet, 1886: 382). A third species, P. potaninii Kom., was described in 1921 based on a specimen from the Yalong Valley in West Sichuan. It has deep maroon-red flowers and was said to differ from P. delavayi in having narrower leaf segments, smaller flowers, paler stamen color, and in the absence of the conspicuous involucre, and from P. lutea in having much narrower leaf segments, smaller flowers, and deep maroon-red flowers. Stern (1931) added the yellow-flowered P. trollioides Stapf ex Stern, based on a specimen from Baima Shan, Deqen County, Northwest Yunnan. In addition, a number of infraspecific taxa have been described, including P. delavayi var. atropurpurea Schipcz., P. delavayi var. angustiloba Rehder & E. H. Wilson, P. delavayi var. alba Bean, and P. delavayi var. lutea f. superba Lemoine.
Paeonia lutea var. ludlowii Stern & G. Taylor, described in 1953 from Southeast Xizang (Tibet), has recently been shown by Hong (1997) to be quite different from the remaining taxa of Paeonia lutea in a number of characters. It is now recognized as a distinct species, P. ludlowii (Stern & G. Taylor) D. Y. Hong, and has been excluded from the P. delavayi complex under study.
Finet and Gagnepain (1904) treated Paeonia lutea as P. delavayi var. lutea (Delavay ex Franch.) Finet & Gagnep. In his monograph of Paeonia, Stern (1946) recognized in this complex three species (P. delavayi, P. lutea, and P. potaninii) and reduced P. trollioides to a variety of P. potaninii. Fang (1958) followed Stern (1946) in the treatment of this complex, but ignored P. lutea var. ludlowii. Wu (1984) recognized two species, P. delavayi, P. delavayi var. angustiloba (= P. potaninii) and P. lutea. Gong (1990) also followed Stern (1946) and recognized three with two infraspecific taxa (P. delavayi, P. lutea, P. potaninii, P. potaninii var. trollioides, and P. potaninii f. alba). However, Pan (1979, 1993) recognized only P. delavayi, with two varieties, var. lutea and var. angustiloba and ignored (Pan, 1979) P. lutea var. ludlowii. Obviously, there is a lack of agreement among the taxonomists who have studied this complex. They have variously emphasized the presence of a conspicuous involucre (P. delavayi), petal color (dark red or deep maroon-red in P. delavayi and P. potaninii, yellow in P. lutea and P. potaninii var. trollioides, white in P. potaninii f. alba), and the width of leaf segments (narrower in P. potaninii than in P. delavayi and P. lutea). The aim of the present paper is to present a revision of this fascinating group of woody peonies based on extensive field observations, population sampling, and analysis of certain characters.
Materials and Methods
In order to fully understand the variation of certain characters, especially flower color, presence of an involucre, and width of leaf segments, the first author (alone or with the other two) made five trips to the distribution range of this group. The first (in 1988) and second (1993) were to the West Hill of Kunming; the third to West Sichuan (1995), including Yajiang County, the type locality of both P. potaninii and P. delavayi var. angustiloba; the fourth to Southeast Xizang (1996), including the type locality of P. lutea var. ludlowii; and the fifth to Northwest Yunnan (1997), including the type localities of P. delavayi, P. lutea, and P. trollioides. In addition to the critical examination of all morphological characters and their variation, biological features (e.g., seed set, presence or absence of cloning) were also recorded.
Eighteen populations were studied in the field by the first author (alone or with the other two), and vouchers are deposited in PE, A, CAS, K, MO, and US. The populations studied are vouchered by the following herbarium specimens (collection numbers of Hong): H97077, H97078, H97087, H97095, H97102, H97103, H97108, H97112, H97119, and H97128 (from Yunnan); H95063, H95070, H95074, and H97110 (from Sichuan); and H96003, H96019, H96024, and H96028. In addition, hundreds of other herbarium specimens from CPB, E, KUN, PE, WU, and XZ were studied morphologically.
Observations and Discussion
Plants of the Paeonia delavayi complex are always dwarf shrubs. The tallest plants (ca. 1.8 m) were found in Yunshanping in Lijiang, Northwest Yunnan (population H97103), where they grew in Picea likiangensis forest at an altitude of ca. 3200 m. By contrast, the shortest plants (rarely reaching 1 m) were found in Ganghaizi, Lijiang (H97095), a population only about 20 km southeast of the first population, and grew in dry, sparse Pinus densata--Quercus gilliana forest. Many short individuals had underground woody parts, and only current-season shoots were aboveground. Plants of the other populations that we studied were intermediate between these two.
Vegetative reproduction is probably predominant in the Paeonia delavayi complex, and seedlings are very rarely found in the field. It is even more predominant at the northwestern and northern boundaries of the distribution range. In Yajiang County, West Sichuan, a population (H95070) was found near a village, where it was represented by some individuals growing by fences and on newly stabilized debris. In this population, only about 50% of the follicles were developed and, because of insect damage, only 20% had seeds. Cloning by stolons, however, was common. Based on an examination of all follicles in the spring of 1996, the five populations observed in Tibet in 1995 produced no seeds. No follicles were observed in a population (temple ruins in Xituan village of the Gengzhanglungba valley, Nyingchi County, Tibet) that consisted of numerous individuals and occupied an area of about 250 m2. This "population" probably consisted of individuals produced through cloning. Cloning was found in every population visited (Figs. 1, 2). The production of fusiform, thickened roots is characteristic of the Paeonia delavayi complex (Fig. 2).
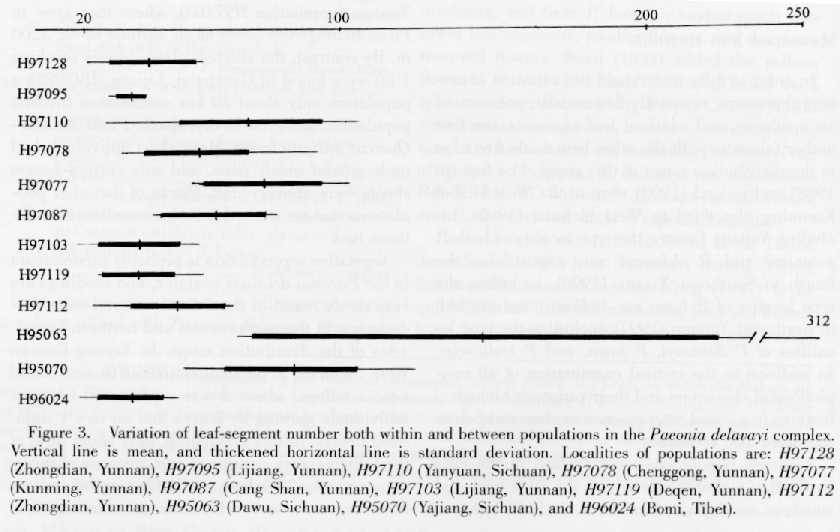
Although leaves in the Paeonia delavayi complex are always biternate, the leaf segments are quite variable in number, length, and width. The number of segments ranged widely from 17 to 312 (Fig. 3), and seemed to differ between populations. For example, the number varied from 17 to 49 in population H96024, and from 68 to 312 in population H95063. However, the standard deviation of the variation in leaf segment number (Fig. 3) shows that these two populations were just the extremes of a wide variation that was also observed within given populations. Taking all the populations into consideration, the number of segments varied continuously in the complex.
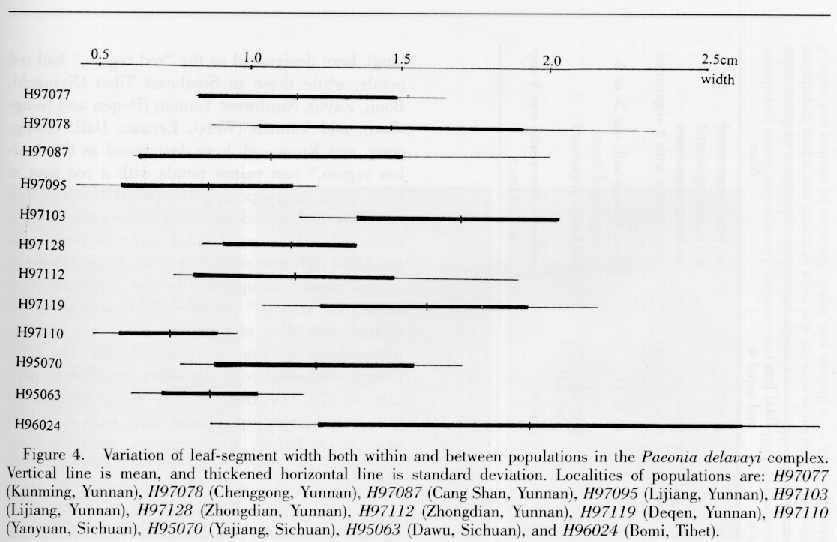
Paeonia potaninii was described as new because it was considered to have narrower leaf segments than P. delavayi and P. lutea. However, we observed that the width of leaf segments varies greatly, and ranges from 0.4 to 2.86 cm within the complex, and from 0.76 to 1.83 cm in population H95070, the type locality of P. potaninii, which falls in the middle of the overall variation range in the complex (Fig. 4). The same situation was found for the length of leaf segments. Therefore, it is evident that P. potaninii is rather similar to P. delavayi and P. lutea in this regard.
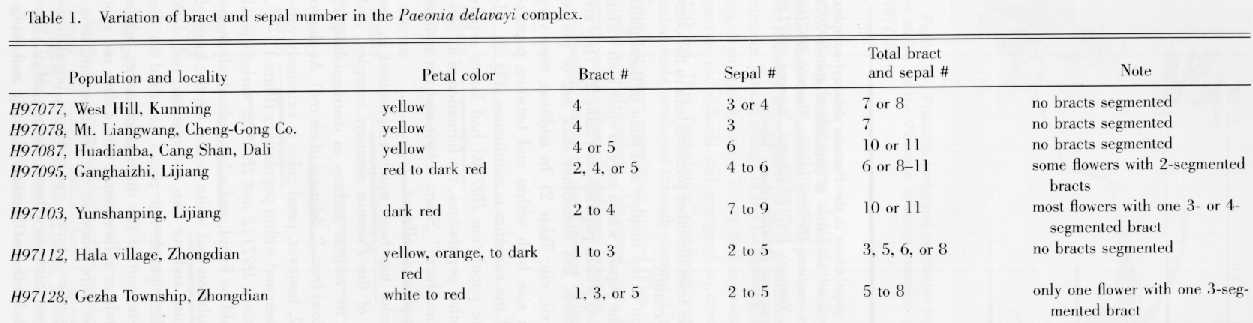
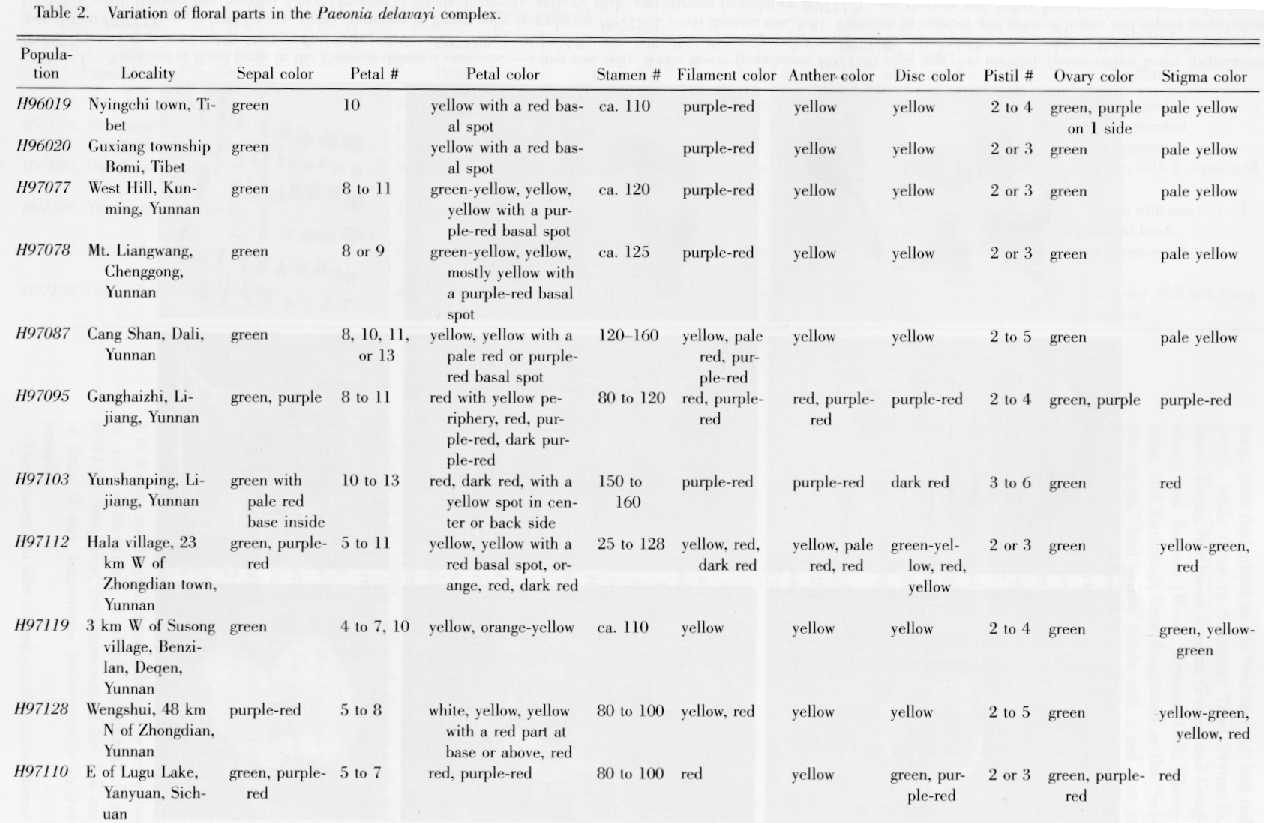
Stern (1946) distinguished Paeonia delavayi within the complex by the presence of a conspicuous involucre immediately below the calyx. However, in the whole genus it is difficult to distinguish clearly between the bracts and sepals, and there is a gradation between leaves and bracts as well. We designate the laminae that are borne some distance below the flowers as leaves, while those at the top of shoots and immediately below the calyx are bracts. Bracts, so designated, have various forms, ranging from segmented and leaflike to entire and sepal-like. The sepals have a much broader proximal part and a dark green, smaller, and narrower distal part with a mucronate or rounded apex. The total number of bracts and sepals varied greatly both within and between populations of the Paeonia delavayi complex. Population H97103 (with dark red flowers) at Yunshanping, Lijiang, the type locality of P. delavayi (Stern, 1946), indeed had the highest number (10 or 11) bracts and sepals forming the so-called "conspicuous involucre," while the other populations observed had fewer bracts and sepals. However, the difference was not distinct (Table 1). Population H97095, only about 20 km from H97103, was very variable in the number of both bracts and sepals, and some flowers had 10 or 11 bracts and sepals (Plate 1, vii), just as those in population H97103. Population H97087 had the same total number of bracts and sepals, and its petals were pure yellow or yellow with a dark red spot at the base. Table 1 and the remarks above show clearly that there is continuous variation in the number of bracts and sepals, and no correlation between petal color and the total number of bracts and sepals. Therefore, P. delavayi is not distinct from the other putative taxa in the complex under study.
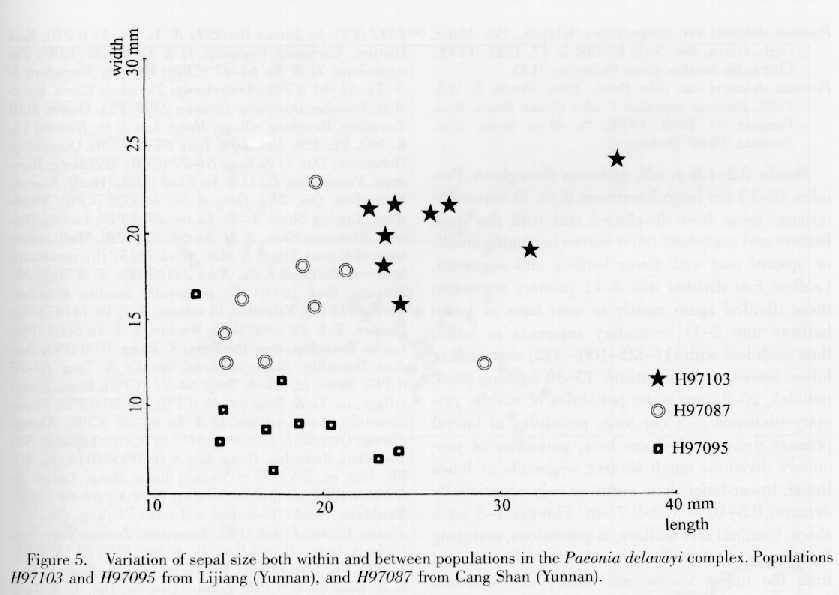
Variation in the number and color of floral parts involves the sepals, which vary in color both within a flower and within populations (Table 2). They are usually green, but sometimes dark red or purple (Plate 1, vii, viii; Table 2). In addition, they vary greatly in size both within and between populations, and the variation is continuous (Fig. 5). Although the population H97103 had larger sepals than the other populations, the formation of a "conspicuous involucre" (Stern, 1946) was not unique in the complex, as alleged.
Petal color has been much emphasized in the taxonomy of the Paeonia delavayi complex, and was used by various authors in distinguishing P. lutea (yellow) from P. delavayi (dark red). As shown in Table 2, however, petal color is extremely variable between and within populations (Plate 1, i--vi). In populations H97112 and H97128, various petal colors could be found, and a few individuals in the latter population had white petals. On the basis of the literature and our own observations, red, dark red, or dark purple-red petals occurred in the northeastern part of the distribution range, while yellow petals or yellow petals with a dark red spot at the base were found in the northwestern, western, and southern parts. For example, populations in Southwest Sichuan (Dawu, Yajiang, Muli, Yanyuan) and Northwest Yunnan (Lijiang and Ninglang), here designated as the "red region," had red petals, while those in Southeast Tibet (Nyingchi, Bomi, Zayu), Northwest Yunnan (Deqen and Gongshan), and Yunnan (Weixi, Eryuan, Dali, Chenggong, and Kunming), here designated as the "yellow region," had yellow petals with a red spot at base, pure yellow petals, and petals either yellow or yellow with a dark red spot at the base, respectively. Therefore, petal color shows a weak geographical differentiation, but it would be unwise to accord formal recognition to the populations in these two "regions" because petal color is uncorrelated with other characters and is very variable within a given "region" or population. The most pronounced petal-color variation occurred in populations H97119 and H97078 of the "yellow region," and some specimens with yellow flowers (e.g., Rock 16110, Rock 161576, Kingdon-Ward 4043, Forrest 12565, and McLaren 89, all at E) were also found within the "red region." Furthermore, petal color was extremely variable in populations such as H97112, H97128, both from Northwest Yunnan.
Petal number in the Paeonia delavayi complex varied enormously, and ranged from 4 to 13, but it also varied greatly within populations. For example, petals were 4--7 or 10 in population H97119, and 8, 10, 11, or 13 in H97087.
Stamen number and color were also very variable. In population H97112 the number varied from 25 to 128. The filaments were yellow, pale red, red, or dark red, and the anthers were yellow, orange, red, or purple in populations H97077, H97078, H97087, H97095, H97103, H97110, and H97112 (Plate 1, viii). But in the populations H97119 and H97128, both filaments and anthers were yellow even in flowers with red petals. The color of the filaments and anthers was not always correlated with that of the petals, although flowers with red petals usually had red or purple filaments and anthers. We conclude that the characters of the androecium cannot be used for dividing this complex.
Unlike the nectar disk of the other woody peonies of section Moutan, the disk in the Paeonia delavayi complex is generally short and fleshy. In populations H97112 and H97119, the disk secreted abundant nectar in some flowers, and it seems likely that the secretion made these flowers more scented. The disk (including teeth) varied in height from 1 to 3 mm, and in color from pale yellow or yellow to red, even within a single population (e.g., H97112). Therefore, the characters of the disk are of little, if any, importance taxonomically in this complex.
The gynoecium also varied greatly in the Paeonia delavayi complex. Pistil (carpel) number was usually 2--4 in most of the populations studied, but in population H97103 it was mostly 5 and very rarely 3, 4, or even 6, and in one collection, Dai, Li & Tang 64-4 (CPB), it ranged from 4 to 8. This variation in pistil number was found in every population observed (Plate 1, ix; Table 2). The pistils were usually green, with a pale green, yellow, or red stigma, but some individuals in populations H97095, H97103, H97112, H97110, and H97128 had entirely purple pistils (Plate 1, viii, ix). Although characters of the gynoecium are generally regarded as significant in the taxonomy of angiosperms, our observations show that they are insignificant taxonomically in this complex.
In summary, the Paeonia delavayi complex exhibits tremendous and continuous variation in characters of the leaves (number, length, and width of segments), bracts (number), and floral parts (color and number). Except for the very weak correlation between petal color and geography, the variation in these characters is clearly insignificant taxonomically. Therefore, none of these characters can be considered as justifying the subdivision of this complex, which we thus consider as comprising only one species, without infraspecific taxa.
Taxonomic Treatment
Paeonia delavayi Franch., Bull. Soc. Bot. France 33: 382. 1886. TYPE: China. NW Yunnan: Lijiang (Likiang), Delavay 1142 (holotype, P; isotype, K).
Paeonia lutea Delavay ex Franch., Bull. Soc. Bot. France 33: 382. 1886. Paeonia delavayi var. lutea (Delavay ex Franch.) Finet & Gagnep., Bull. Soc. Bot. France 51: 524. 1904. TYPE: China. NW Yunnan: Eryuan County, Mt. Hea Chan Men, 25 May 1883, Delavay s.n. (lectotype, here designated, P; isolectotype, K).
Paeonia potaninii Kom., Bot. Mater. Gerb. Glavn. Bot. Sada RSFSR 2: 7. 1921. TYPE: China. W Sichuan: Yajiang (Tachien-lu district in Yalung valley), Potanin s.n. (holotype, LE).
Paeonia trollioides Stapf ex Stern, J. Roy. Hort. Soc. 56: 77. 1931. Paeonia potaninii var. trollioides (Stapf ex Stern) Stern, Stud. Paeonia 50. 1946. TYPE: China. NW Yunnan: Deqen, Baima Shan, Mekong-Yangtse divide, 11,000 ft., open stony pastures, Forrest 13195 (holotype, E).
Paeonia delavayi var. lutea f. superba Lemoine, Rev. Hort. 14: cum tab., 1906. Paeonia lutea var. superba (Lemoine) hort. ex Gard. Chron., Ser. 3. 44: 50, cum tab., 1908. TYPE: ["pl. 14 in Lemoine, Rev. Hort. 1906"].
Paeonia delavayi var. angustiloba Rehder & E. H. Wilson, in Sarg., Pl. Wilson. 1: 318. 1913. TYPE: China. W Sichuan: Yajiang (W of Tachien-lu, descent to Yalung river), 3000--3600 m, Oct. 1908, Wilson 1333 (holotype, A).
Paeonia delavayi var. atropurpurea Schipcz., Bot. Mater. Gerb. Glavn. Bot. Sada RSFSR 2: 47. 1921. TYPE: China. No locality given (holotype, ?LE).
Paeonia delavayi var. alba Bean, Trees Shrubs 3: 265. 1933. Paeonia potaninii f. alba (Bean) Stern, Stud. Paeonia 49. 1946. TYPE: "t. 49 in Stern, Stud. Paeonia, 1946" (holotype).
Shrubs 0.2--1.8 m tall, glabrous throughout. Petioles 10--15 cm long; lowermost 2 (or 3) leaves biternate, these best developed and with the most leaflets and segments, other leaves becoming smaller upward and with fewer leaflets and segments. Leaflets first divided into 3--11 primary segments, these divided again mostly to near base or lobed halfway into 2--11 secondary segments or lobes, thus each leaflet with (17--)25--100(--312) segments or lobes; leaves ovate in outline, 15--30 cm long (excl. petiole), 10--22 cm wide; petiolules of middle primary divisions 5--9 cm long, petiolules of lateral primary divisions 1--3 cm long, petiolules of secondary divisions much shorter; segments or lobes linear, linear-lanceolate, entire or only occasionally dentate, 0.5--10 ´ 0.2--2.7 cm. Flowers 1--3 on a shoot, terminal and axillary, ± pendulous, maturing basipetally. Bracts 1--5, gradually differentiated from the upper leaves and hardly distinguished from the sepals, the outer ones 2--4-segmented, green. Sepals 2--9, green outside, green with a pink base inside, or entirely purple or purple-red, rounded or triangular-rounded, acuminate or mucronate to rounded at apex, 1.3--3.7 ´ 0.6--2.3 cm. Petals (4--)7--11(--13), yellow, yellow with a red or purple-red basal spot, red, dark red, or dark purple-red, sometimes white, orange, green-yellow, or yellow with red margin. Stamens 25--160; filaments yellow, pink, red, or dark purple-red; anthers yellow, pink, red, or dark purple-red. Disks short, annular or short-cylindric, 1--3 mm high, dentate, green, yellowish, yellow, red, or dark red. Carpels 2--4, very rarely 6--8; ovary usually green, sometimes purple; stigma yellow-green, yellow, red, or purple-red; ovules 7--17 per carpel. Follicles oblong-ovoid, 2--3.5 ´ 1--1.5 cm, brown at maturity. Seeds 1--6 in each follicle, brown-black.
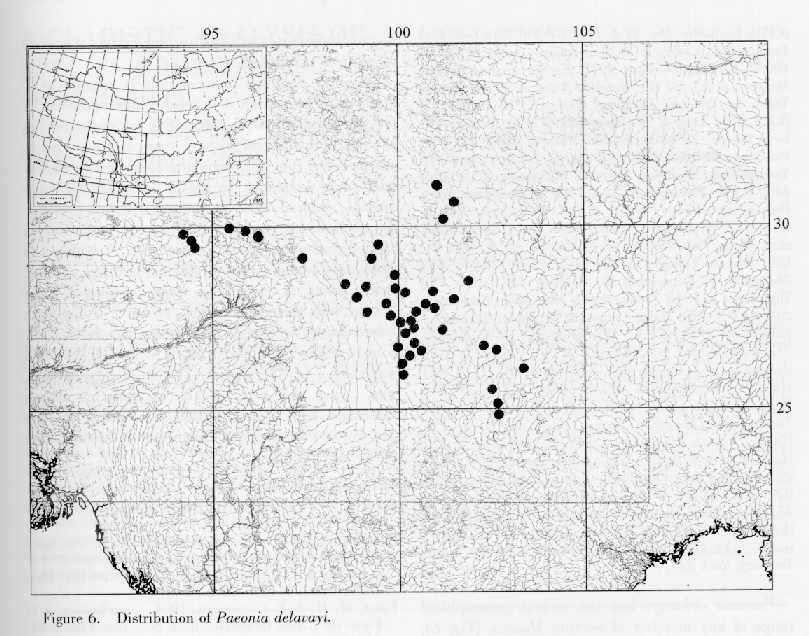
Distribution. Endemic to China and restricted to Sichuan, Tibet (Xizang), and Yunnan. Paeonia delavayi grows at 2000--3600 m, primarily in sparse thickets or dry Pinus and Quercus woods, or rarely in grassy slopes or glades of virgin Picea forests (Fig. 6).
Phenology. Flowering from mid May to mid June; fruit maturing August. 2n = 10 (Gong et al., 1991).
Additional specimens examined (all from CHINA). SICHUAN. Batang: Zongza Township, Anonymous 1547 (PE), by Jinsan River, Li & Xu 64-43 (CPB); East District, Yanrigong Township, Li & Xu 64-38 (CPB); Zangqenrong, Li & Xu 64-47 (CPB); Baisong Township, Li & Xu 64-48 (CPB). Daocheng: Dongnyi, Gawa Township, Kasishe, Daocheng Division 2397 (PE). Dawu: Mazi Township, Benglong village, Hong, Luo & He H95063 (A, K, MO, PE, US); Dai, Li & Tang 64-4 (CPB); Qiangning, Ochancun, Dai, Li & Tang 64-27 (CPB). Huidong: Baisanpo, Paomaping, Li, Li & Xu 64-9 (CPB). Huili: Xiacun, Heilaoling, Qiu, Zhu, Deng & Shi 66-023 (CPB). Mianning: Jinping Shan, Z. M. Xu 64-27 (CPB); Luning District, Xiamatou Shan, Z. M. Xu 64-30 (CPB). Muli: mountains of Kopati, Diago & Muli, Rock 16157 (E); mountains between Muli and Kulu, Rock 24123 (E); W of Muli, Mt. Mitzuga, Rock 16110 (E); no precise locality, Kingdon-Ward 4043 (E); Yalientsa, in valleys, T. T. Yü 14147 (PE); Chaiwu, T. T. Yü 5560 (PE); Wachin, T. T. Yü 6003 (PE); Luobo Township, near the ferry, S. Jiang 5708 (PE); Baidiao Township, Shangjiamiwan, Li, Xu & Tang 64-17 (CPB); Boao, Li, Xu & Tang 64-11 (CPB); Boao, Shuzu village, Li, Xu & Tang 64-19 (CPB), 64-20 (CPB); Zhuao Township, Aoxia village, Li & Xu 64-26 (CPB). Xiangcheng: Qingda, L. D. Sheng 64-113 (CPB). Yajiang: Niri Township, Bajiaolou, Hong, Luo & He H95070 (A, K, MO, PE, US); ca. 5 km E of Yajiang Town, Hong, Luo & He H95074 (PE); no precise locality, Li & Xu 64-64 (CPB). Yanbien: Yanshi Township to Jiudao Zhuling, Qinghai-Xizang Exped. 11458 (PE). Yanyuan: Zuosuo Township, by Lugu Lake, Hong, Pan, Yu & Dai H97110 (A, CAS, K, MO, PE, US); Zuosuo Township, near Shanjiacun, Dai, Li & Tang 64-6 (CPB); Zuosuo District, Dai, Li & Tang 64-1(CPB); Yantang, Dacaoxiang, Nancaowan, Dai, Li & Tang 64-7 (CPB); Shangjiagou, Dai, Li & Tang 64-17 (CPB); E of Yongning (Yunnan), 27°50¢N, 100°56¢E, Forrest 20458 (E); Lumapu, 27°37¢N, Handel-Mazzetti 2067 (E, WU). TIBET (Xizang). Bomi: Guxiang Township, Ying & Hong 39 (PE), 214 (PE), Hong, Luo & Zhang H96024 (A, K, MO, PE, US); between Zamu & Guxiang, Zhang & Lang 379 (PE); near Zamu, behind Army Station, Xiao, Xia & Mi 2233 (KUN, PE); Sumzom Township, Sumzom, S of River, Hong, Luo & Zhang H96028 (A, K, MO, PE, US). Markam: Cawarong, Chaila, C. W. Wang 66335 (PE); Cawarong, Dula, C. W. Wang 65523 (PE); Cawalong, Songta Snow Mountain, Qinghai-Xizang Exped. 7633 (KUN, PE); E Himalayas, no precise locality, Kingdon-Ward 5691 (E). Nyingchi: by Nyingchi Town, Hong, Luo & Zhang H96019 (A, K, MO, PE, US); Gengzhang Nongba Valley, Xituan Village, Hong, Luo & Zhang H96003 (A, K, MO, PE, US); Bayi Town, Jiaomugou, Hong, Luo & Zhang H96004 (A, K, MO, PE, US). Zayu: Gujing District, Xizang Biol. Inst. Pl. Resources Exped. 3895 (XZ), near Guyu to Cinong, Qinghai-Xizang Exped. 73-294 (PE); Gujing District, Ni, Wang, Cidou & Cidan 1501 (PE, XZ); no precise locality, Ludlow & Sherriff 1376 (E).
YUNNAN. Chenggong: Mt. Liangwan, Hong, Pan, Yu & Dai H97078 (A, CAS, K, MO, PE, US). Dali: Mt. Cangshan, Huadianba, Hong, Pan, Yu & Dai H97087 (A, CAS, K, MO, PE, US), R. C. Ching 22954 (KUN, PE), Sino-British Cangshan Exped. 0684 (E, KUN), Xiaohuadianba, Q. Ling 7708 (KUN); no precise locality, T. N. Liou 16149 (KUN, PE), T. N. Liou 20677 (KUN, PE), McLaren 55 (E); E flank of the Cangshan range, Forrest 4352 (E), 6787 (E), 30998 (E); Mt. Pi-iou-se, above Tapin-tze, 11 June 1883, Delavay s.n. (P); Tali (Dali), Mt. Che-tcho-tze, 9 May 1883, Delavay s.n. (P); Tali (Dali), Mt. Che-tcho-tze, 10 Oct. 1883, Delavay s.n. (P). Deqen: Benzilan, 3 km W of Susong village, Hong, Pan, Yu & Dai H97119 (A, CAS, K, MO, PE, US); Cizhong to Yongzi, by the Lancang River, K. M. Feng 5765 (KUN, PE); Yunling, Nanshui Beidiao Exped. 9194 (KUN, PE); E flank of Baima Shan, Benzilan, Qinghai-Xizang Exped. 11795 (KUN, PE); Benzilan, Yonglobu (Forest Farm), Qinghai-Xizang Exped. 1878 (KUN, PE); Benzilan, Dongzuling, Qinghai-Xizang Exped. 2209 (KUN, PE); no precise locality, C. W. Wang 70211 (KUN, PE); Baima Shan, Rock 22821 (E). Dongchuan: Jinniu, D. C. Liu 0447 (CPB); Mt. Lo Shivel, McLaren V2 (E). Gongshan: Suroula, C. W. Wang 66617 (KUN, PE); Mekong-Salwin divide, 28°12¢N, Forrest 16339 (E), Forrest 16527 (E). Heqing: Laugkong-Hoching divide, 26°16¢N, Forrest 10062 (E); Baiya, Sanxi, R. C. Ching 24365 (KUN, PE); Songgui, Mt. Maer, R. C. Ching 24191(KUN, PE). Jianchuan: Xinsheng Township, Huajiaping, P. Y. Mao 236 (KUN). Kunming: Shanqing temple to Shitoushan, Liou 20677 (PE); West Hill, Hong, Pan, Yu & Dai H97077 (A, CAS, K, MO, PE, US), B. Y. Qiu 51019 (KUN), B. Y. Qiu 57101 (KUN); Seventh District, Shungxiao Township, B. Y. Qiu 51880 (KUN); Longquan District, Heping Township, S. K. Wu 8 (KUN). Lanping: No. 104 Forest Farm, West Hill of Xiaoyanjing, Hengduan Mt. Exped. 908 (PE). Lijiang: Yulong Snow Range, Ganghaizi, Hong, Pan, Yu & Dai H97095 (A, CAS, K, MO, PE, US), Qinghai-Xizang Exped. 201 (PE), Edinburgh Bot. Gard. Exped. 85-6 (E, KUN), R. C. Ching 20438 (KUN, PE), Z. W. Lu 1002 (KUN), Kingdon-Ward 238 (E); Yulong Snow Range, Muzhugou to Ganghaizi, X. Zhou 1030 (KUN), Qingmugou to Ganghaizi, Lijiang Bot. Gard. 100486 (KUN); Yulong Snow Range, Yunnan Univ. Biol. Dept. Vegetation Exped. 233 (KUN), R. C. Ching 30987 (KUN, PE); Yulong Snow Range, Yuhu, Xuesong Village, Mt. Beibazi, R. C. Ching 30175 (KUN, PE), K. M. Feng 21245 (KUN), E of the village, R. C. Ching 30071 (KUN, PE); near Yuhu, Ameidiruo, K. M. Feng 22229 (KUN); Yulong Snow Range, Wutoudi, K. M. Feng 23029 (KUN, PE); Yulong Snow Range, Baishui, Hong, Pan, Yu & Dai H97102 (A, CAS, K, MO, PE, US); Yulong Snow Range, Yunshanping, Hong, Pan, Yu & Dai H97103 (A, CAS, K, MI, PE, US); Yulong Snow Range, Mahuangba to Wutoudi, Z. H. Yang & Y. C. Cai 101777 (PE); Yulong Snow Range, Jiuzihai to Yulong Snow Range, Heishui, J. S. Yang 4157 (KUN); Heishui River, left slope, Lijiang Bot. Gard. 100026 (KUN); Yulong Snow Range, Mt. Gyina Loko, Rock 24984 (E); E flank of Yulong Snow Range, Forrest 5716 (E), T. T. Yü 15016 (E, PE); Mt. Wenbi, T. T. Yü 8107 (KUN, PE); northern Part, Hongshiyan, R. C. Ching 20595 (KUN, PE); Shapingze, T. T. Yü 5160 (KUN, PE), T. T. Yü 5163 (KUN, PE); Sixth District, X. L. Wang s.n. (CPB); Zili, by the river, R. C. Ching 22192 (KUN, PE); S of Lijiang, Sungkwe Pass between Lijiang & Heqing, Rock 25179 (E); 22 km from Lijiang to Yongsheng, Hong, Pan, Yu & Dai H97108 (PE). Luquan: Xinming Township, Laoyingya, P. Y. Mao 939 (KUN); Emao Township, Mt. Dangan, P. Y. Mao 01565 (KUN). Ninglang: Gouzuandong, J. S. Yang 4123 (KUN); Yongning, Geao Pass, J. S. Yang 4130 (KUN); hills around Yongning, Forrest 12503 (E), 12565 (E); Yongning, Shize Shan, T. T. Yü (PE); Yongning, Wenquan Township, near Wenquan, Dai, Li & Tang 64-2 (CPB), Liujiashe, Dai, Li & Tang 64-3 (CPB); Dai, Li & Tang 64-4 (CPB); E of the Yangtze bend, 27°35¢N, 100°45¢E, Kingdon-Ward 3981 (E); no precise locality, McLaren 5 (E), 89 (E), Kingdon-Ward 5055 (E). Weixi: Yezhi, by Lancang (Mekong) River, K. M. Feng 4220 (KUN, PE); Yezi, C. W. Wang 68199 (PE); Yangtze-Mekong divide, 29°45¢N, Handel-Mazzetti 8868 (E, WU); Pantiange District, Wucun village, D. Y. Chen s.n. (CPB). Yongsheng: Shunzhou District, Shisanzuo, J. S. Yang 4402 (CPB). Zhongdian: 23 km NW of Zhongdian Town, Hala village, Hong, Pan, Yu & Dai H97112 (A, CAS, K, MO, PE, US); 45 km N of Zhongdian Town, Gezan Township, Hong, Pan, Yu, Dai H97128 (A, CAS, K, MO, PE, US); Wucun, Gongbi, K. M. Feng 23561 (KUN, PE); Zhongdian to Annachang, K. M. Feng 938 (KUN, PE); Tuguancun to Haba Xuemenkang, Zhongdian Exped. 63-2389 (KUN, PE); Haba, Longwang Binsanba, Zhongdian Exped. 63-2738 (KUN, PE); Huangdong, Reshuitang, Zhongdian Exped. 63-2597 (KUN, PE); Qiaotou to Xiaozhongdian, near Hongtu xiao, K. M. Feng 885 (KUN, PE); Haba to Xiaozhongdian, K. M. Feng 1387 (KUN, PE); Baidi, T. T. Yü 11389 (KUN, PE); T. T. Yü 14915 (PE); Tuguancun, Zhongdian Exped. 63-2375 (KUN, PE); Haba, Annazai, Zhongdian Exped. 63-2647 (KUN, PE); Haba, near Longwanbin, Zhongdian Exped. 1634 (KUN, PE); SE of Zhongdian Town, Jiuluo, Zhongdian Exped. 929 (KUN, PE); no precise locality, 27°50¢N, Forrest 15162 (E); Haba Shan, N of the Yangtze loop, Rock 24758 (E); no precise locality, 27°30¢N, Forrest 12561(E); Chihren, T. T. Yü 11247 (E, PE); N of Zhongdian Town, Tonwa Territory, Rock 24717 (E).
Paeonia delavayi has the widest geographical range of any member of section Moutan (Fig. 6). The plants reproduce predominantly vegetatively, and cloning by stolons (Fig. 1) was commonly seen in every population visited except for population H97103, in Picea likiangensis forest in Yunshanping Lijiang, Yunnan. Vegetative reproduction is particularly prominent in West Sichuan (Dawu, Yajiang) and Southeast Tibet, where seedlings have never been found and clones often covered large areas. The roots of P. delavayi are always fusiformly thickened (Fig. 2). Such roots and stolons probably make the species more adapted to open, somewhat dry and disturbed habitats, and enable the species to become established very rapidly in a given population. It may also account for the scattered distribution of the species and the large number of "individuals" in a given population. Paeonia delavayi has been listed as an endangered species in the China Plant Red Data Book (Feng, 1992). On the basis of its vegetative reproduction and relatively wide distribution, however, it is reasonable to conclude that it will not become extinct if wanton digging is controlled.
Paeonia delavayi is most closely related to P. ludlowii, from which it is readily distinguished by having predominantly vegetative reproduction by stolons and fusiform roots, a non-caespitose habit, stems 0.2--1.8 m tall, segmented leaves with narrow and acute segments, variously colored floral parts (petals, stamens, disk, and stigmas), a 2--8-pistillate gynoecium, and small (2--3.5 ´ 1--1.5 cm) follicles rarely producing seeds. In contrast, P. ludlowii is obligately sexual, without stolons, and has slender, terete roots, a caespitose base forming a large clump with dozens of stems, stems 2--3.5 m tall, lobed leaves with short and acuminate lobes, uniformly yellow floral parts (petals, stamens, disk, and stigmas), a 1(or very rarely 2)-pistillate gynoecium, and larger (4.7--7 ´ 2--3.3 cm) follicles always bearing seeds. In fact, P. delavayi and P. ludlowii are better considered as members of two separate species complexes.
Literature Cited
Fang, W.-P. 1958. Notes on Chinese paeonies. Acta Phytotax. Sin. 7: 297--323.
Feng, K.-M. 1992. Paeonia delavayi Franch. var. lutea (Delavayi ex Franch.) Finet & Gagnep. Pp. 530--531 in L.-K. Fu & J. M. Jin (editors), China Plant Red Data Book, Vol. 1. Science Press, Beijing.
Finet, M. M. & F. Gagnepain. 1904. Contributions à la Flore de L’Asie Orientale. Bull. Soc. Bot. France 51: 524--525.
Franchet, A. 1886. Plantas yunnanensis. Bull. Soc. Bot. France 33: 358--467.
Gong, X. 1990. A Taxonomical Study of the Paeonia delavayi complex. Master’s Thesis, Institute of Botany, Beijing.
Gong, X., Z. J. Gu & Q. A. Wu. 1991. A cytological study of seven populations in Paeonia delavayi var. lutea. Acta Bot. Yunnan. 13: 402--410.
Hong, D.-Y. 1997. Paeonia (Paeoniaceae) in Xizang (Tibet). Novon 7: 156--161.
Pan, K.-Y. 1979. Paeonia. In: T.-W. Wang (editor), Fl. Reipubl. Popularis Sin. 27: 37--59. Science Press, Beijing.
---. 1993. Paeoniaceae. Pp. 545--546 in W.-T. Wang & S.-G. Wu (editors), Vascular Plants of the Hengduan Mountains. Science Press, Beijing.
Stern, F. C. 1931. Paeony species. J. Roy. Hort. Soc. 56: 71--77.
---. 1946. A Study of the Genus Paeonia. Royal Horticultural Society, London.
--- & G. Taylor. 1953. Paeonia lutea var. ludlowii. Bot. Mag. 169: tab. 209.
Wu, C. Y. (Editor). 1984. Index florae yunnanensis. 1: 122--123. The People’s Publishing House, Kunming, China.
Fieldwork was financially supported by the National Geographic Society (Grant 5515-95), to which the authors are very grateful. The authors are indebted to Luo Yi-bo and Zhang Shu-ren (Institute of Botany, Beijing) and He Yong-hua (Chengdu Institute of Biology, Chinese Academy of Sciences) for their assistance during fieldwork. We thank Li Qiao-ling for the measurements and calculations in the statistical study, and for typing the manuscript. Our sincere gratitude is due to Ihsan Al-Shehbaz for his encouragement and help with the manuscript. Laboratory of Systematic and Evolutionary Botany, Institute of Botany, Chinese Academy of Sciences, 20 Nanxincun, Xiangshan, Beijing 100093, People’s Republic of China.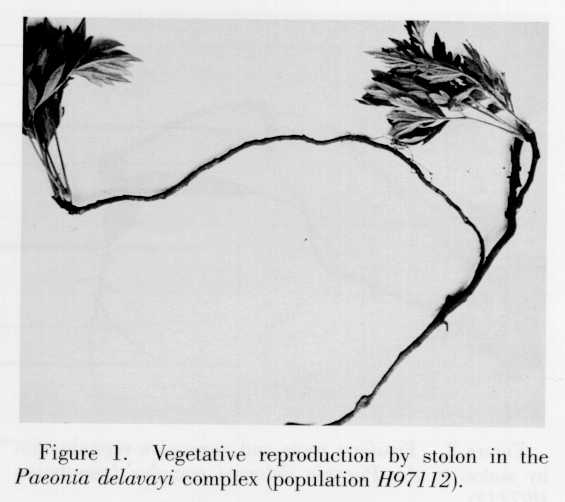{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}